Ich charakteristikou sú vestibulárne reflexy. Vestibulárne dráhy a reflexy. Štruktúra dráh a centier vestibulárneho systému
Neuróny vestibulárnych jadier zabezpečujú kontrolu a riadenie rôznych motorických reakcií. Vestibulospinálne vplyvy menia impulzy neurónov na segmentových úrovniach miechy. Takto prebieha dynamická redistribúcia tonusu kostrového svalstva a zapínajú sa reflexné reakcie potrebné na udržanie rovnováhy. Podieľa sa na vestibulo-vegetatívnych reakciách kardiovaskulárneho systému, gastrointestinálny trakt a ďalšie vnútorné orgány. Pri silnom a dlhotrvajúcom zaťažení vestibulárneho aparátu sa vyskytuje pohybová choroba (napríklad morská choroba). Vestibulo-okulomotorické reflexy (očný nystagmus) spočívajú v pomalom rytmickom pohybe očí v opačnom smere k rotácii, po ktorom nasleduje ich skok späť. Výskyt a charakteristika rotačného očného nystagmu sú dôležitými indikátormi stavu vestibulárneho systému a sú široko používané v experimentoch a klinikách.
Koniec práce -
Táto téma patrí:
Základy psychofyziológie: Učebnica / Ed. vyd. Yu.I. Alexandrov
Na stránke si prečítajte: 075 základy psychofyziológie: učebnica / resp. vyd. y.i. alexandrov. - m.: infra-m, 1997. - 349 s...
Ak potrebujete ďalší materiál k tejto téme, alebo ste nenašli to, čo ste hľadali, odporúčame použiť vyhľadávanie v našej databáze prác:
Čo urobíme s prijatým materiálom:
Ak sa tento materiál ukázal byť pre vás užitočný, môžete si ho uložiť na svoju stránku v sociálnych sieťach:
| pípanie |
Všetky témy v tejto sekcii:
Všeobecné informácie
Tradične, od čias francúzskeho fyziológa Bisha (začiatok 19. storočia), sa nervový systém delí na somatický a autonómny, pričom každý z nich zahŕňa štruktúry hlavy a chrbtice.
Neuron. Jeho štruktúra a funkcie
Ľudský mozog pozostáva z 1012 nervových buniek. Bežná nervová bunka prijíma informácie od stoviek a tisícok iných buniek a odovzdáva ich stovkám a tisíckam, a počet spojení
Veľkosť a tvar
Veľkosti neurónov sa môžu meniť od 1 (veľkosť fotoreceptora) do 1000 µm (veľkosť obrovského neurónu v morskom mäkkýšovi Aplysia) (pozri (Sakharov, 1992)). Výnimočný je aj tvar neurónov
Farba neurónov
Ďalšou vonkajšou charakteristikou nervových buniek je ich farba. Je tiež pestrá a môže naznačovať funkciu bunky – napríklad neuroendokrinné bunky sú biele. žltá
synapsie
Biofyzikálny a bunkový biologický prístup k analýze neuronálnych funkcií, možnosť identifikácie a klonovania génov nevyhnutných pre signalizáciu, odhalil úzky vzťah medzi
elektrická excitabilita
Všetky funkcie vlastné nervovému systému sú spojené s prítomnosťou štrukturálnych a funkčných vlastností v nervových bunkách, ktoré poskytujú možnosť tvorby pod vplyvom vonkajších
Kardiostimulátor
Jedným z úžasných typov elektrickej aktivity neurónov zaznamenaných intracelulárnou mikroelektródou sú potenciály kardiostimulátora. A. Arvanitaki a N. Halazonitis. Chemoreceptory informujú CNS o zmenách chemizmu
Dráhy a centrá viscerálneho zmyslového systému
Dráhy a centrá viscerálneho zmyslového systému sú zastúpené najmä vagusovými, celiakálnymi a panvovými nervami. Nervus vagus prenáša aferentné signály do CNS cez tenké
Viscerálny pocit a vnímanie
Excitácia niektorých interoceptorov vedie k vzniku jasných lokalizovaných vnemov, t.j. k vnímaniu (napríklad keď sú steny močového mechúra alebo konečníka natiahnuté). IN
Hlavné kvantitatívne charakteristiky zmyslových systémov človeka
Blízky bod jasného videnia 10 cm Priemer makuly sietnice cca.
Ovládanie pohybu
Pohyb (vrátane reči a písania) je hlavným prostriedkom interakcie ľudského tela s prostredím. V tejto interakcii sa reflexné reakcie, vyvolané environmentálnymi stimulmi, s
Všeobecné informácie o nervovosvalovom systéme
Nie je možné pochopiť princípy fungovania riadiaceho systému bez znalosti štrukturálnych vlastností riadiaceho objektu. S ohľadom na pohyby zvierat a ľudí je predmetom kontroly op
propriocepcia
Pre úspešnú realizáciu pohybov je potrebné, aby centrá, ktoré tieto pohyby riadia, mali kedykoľvek informácie o polohe väzieb tela v priestore a o
Centrálne zariadenia na riadenie pohybu
Na riadení pohybov sa podieľajú takmer všetky časti centrálneho nervového systému – od miechy až po mozgovú kôru. U zvierat môže miecha vykonávať pomerne rozsiahlu triedu
Motorické programy
Ovládanie pohybu je nemysliteľné bez koordinácie činností Vysoké číslo svaly. Povaha tejto koordinácie závisí od motorickej úlohy. Takže, ak si potrebujete dať pohár vody,
Koordinácia pohybu
Koncept koordinácie pohybov vznikol na základe pozorovaní pacientov, ktorí z rôznych dôvodov nie sú schopní plynulo a presne vykonávať pohyby, ktoré sú pre zdravých ľudí ľahko dostupné.
Typy pohybu
Ľudské pohyby sú veľmi rôznorodé, no všetku túto rôznorodosť možno zredukovať na malý počet základných typov činností: zabezpečenie držania tela a rovnováhy, pohyb a svojvôľa.
Rozvoj pohybových schopností
K zlepšeniu motorickej funkcie v ontogenéze dochádza jednak v dôsledku dozrievania vrodených mechanizmov zapojených do koordinácie
Psychofyziológia pamäti
Na učenie sa možno pozerať ako na postupnosť zložitých procesov zapojených do získavania, uchovávania a reprodukcie informácií (pozri kapitolu 15). V dôsledku učenia
Časová organizácia pamäte
Časová organizácia pamäťovej stopy implikuje postupnosť vývoja v čase kvalitatívne odlišných procesov vedúcich k fixácii nadobudnutých skúseností. Základné pojmy, do
retrográdny gradient amnézie
Závislosť účinnosti modulácie pamäte od časového intervalu medzi tréningom a použitím amnestického činidla charakterizuje gradient retrográdnej amnézie. krupobitie
Fázy odovzdania pamäte
Hypotéza dvoch postupne sa rozvíjajúcich stôp. Podľa hypotézy sa tvorba engramu uskutočňuje v dvoch fázach: prvá je charakterizovaná nestabilnou formou
Krátkodobá a dlhodobá pamäť
Rozvoju koncepcie časovej organizácie pamäti bráni neostrosť a vágnosť základných pojmov, na ktorých je založená teoretická konštrukcia. Koncept stručného
Spontánna obnova pamäte
Fakty o spontánnom obnovení pamäti po pôsobení amnestického elektrického šoku boli známe už v 50. rokoch minulého storočia. (pozri v [Grechenko, 1979]). Obnovenie zručností bolo hlásené po
Obnovenie engramu s efektom druhého šoku
Mnohí výskumníci informovali o obnovujúcom účinku sekundárne prezentovanej kombinácie „trest (podnet, ktorý sa používa pri učení) – elektrický šok“ (pozri v)
Obnova pamäte metódou pripomenutia
V diele R.J. Koppenaal a spol., ako prví aplikovali metódu „pripomenutia“. Spočíva v tom, že pred testovaním zachovania zručnosti sa zvieratám predložia
Obnova pamäte oboznámením sa
Ak sa zviera pred výcvikom umiestni do experimentálnej komory a nechá sa v nej voľne pohybovať, potom po aplikácii amnestického činidla retrográdna amnézia nenastane
Retrográdna amnézia na reaktivované pamäťové stopy
Zistilo sa, že po prechode pamäte do dlhodobého skladovania možno vyvolať retrográdnu amnéziu. V experimentoch sme študovali vplyv elektrického šoku na zručnosť vytvorenú pred niekoľkými dňami.
Hlavné postavenie teórie aktívnej pamäte
Hlavné ustanovenia konceptu aktívnej pamäte sú nasledovné. Pamäť pôsobí ako jedna vlastnosť, t.j. neexistuje rozdelenie na krátkodobé a dlhodobé
Hypotéza distribúcie Engramu
Experimenty s lokálnou mozgovou stimuláciou ukázali, že vznik retrográdnej amnézie pri stimulácii určitej štruktúry závisí od časového intervalu, ktorý uplynul od okamihu jej ukončenia.
Distribúcia engramu v experimentoch s lokálnou stimuláciou mozgu
Štúdie vykonané pomocou elektrických výbojov, ktoré spôsobujú rozvoj elektrickej kŕčovej aktivity, ukazujú zložitú dynamiku pohybov aktívneho engramu pozdĺž
Distribúcia engramu cez súbor prvkov mozgu
Myšlienka, že pamäťová stopa nemá špecifickú lokalizáciu, ale je čítaná z neurónov rôznych mozgových štruktúr, v závislosti od okolností, bola potvrdená experimentmi. F
Procedurálna a deklaratívna pamäť
V poslednej dobe sa pojem viacerých pamäťových systémov stáva čoraz dôležitejším. Tento názor bol vytvorený na základe údajov získaných v štúdii
Molekulárne mechanizmy pamäti
V neurovede sa mechanizmy učenia a pamäte študujú predovšetkým v kontexte plasticity (pozri kapitolu 15). Preto sa mnohé štúdie zameriavali na identifikáciu
Diskrétnosť mnemotechnických procesov
Písmená abecedy, atómy a molekuly sú kódy pre dôležité entity, ktorých objavy nemožno preceňovať. Prvým bol objav hieroglyfov a abecedy. Azb
Livanovova konštanta
Existuje mnoho empirických vzťahov nazývaných zákony. Ako príklad môžeme uviesť základný psychofyzikálny zákon, ktorý stanovuje závislosť sily vnemu na f
Veľkosť a rýchlosť pamäte
Ak sa budeme riadiť logikou D. Gartleyho, A.A. Ukhtomsky, N.G. Samojlová, M.N. Livanov, G. Walter, E.R. Jána, K. Příbram a ďalších priaznivcov myšlienky dynamického kódovania vnímame
Rozsah vnemov
V psychofyzike Weberov zlomok definuje hranicu medzi zmyslovým a necitlivým. Ide o sotva badateľný subjektívny nárast vnemov, braný vo vzťahu k počiatočnej hodnote stimulu. vytekať
Kódy neurálnej pamäte
Kódy neurónovej pamäte sú cyklicky sa opakujúce impulzné vlny generované nervovými súbormi. Trvanie jedného cyklu je asi 100 ms. Všimnite si, že podľa údajov
Emócia ako odraz skutočnej potreby a pravdepodobnosť jej uspokojenia
William James, autor jednej z prvých fyziologických teórií emócií, dal svojmu článku publikovanému pred viac ako 100 rokmi veľmi expresívny názov: "Čo je to emócia?" [
Mozgové štruktúry, ktoré implementujú posilňujúce, prepínacie, kompenzačno-substitučné a komunikačné funkcie emócií
Výsledky neurofyziologických experimentov ukazujú, že potreby, motivácie a emócie majú rôzne morfologické substráty. Takže pri stimulácii zón sebapodráždenia neskôr
Jednotlivé znaky interakcie mozgových štruktúr, ktoré implementujú funkcie emócií ako základ temperamentov
Ako metódy na diagnostikovanie stabilných (typologických) individuálnych behaviorálnych charakteristík sme použili dva modely správania: test preferencie pravdepodobnosti alebo hodnotu
Vplyv emócií na aktivitu a objektívne metódy sledovania emočného stavu človeka
Samotný fakt generovania emócií v situácii pragmatickej neistoty predurčuje a vysvetľuje ich adaptívnu kompenzačnú hodnotu. Faktom je, že keď sa objaví emócia
Definícia funkčného stavu
Funkčný stav (FS) je najčastejšie definovaný ako činnosť pozadia nervových centier, v ktorej sa realizuje jedna alebo druhá špecifická ľudská činnosť. V klasickej
Úloha a miesto funkčného stavu v správaní
Funkčné stavy regulované modulačným systémom mozgu sú nevyhnutnou súčasťou akéhokoľvek druhu činnosti a správania. Vzťah medzi úrovňou aktivácie
Stem-thalamo-kortikálny systém
Štúdie J. Moruzziho a G. Maguna ich priviedli k objavu v trupe na úrovni stredného mozgu nešpecifického systému, čiže mezencefalického retikulárneho útvaru, ktorý aktivuje kôru
Bazálny cholinergný systém predného mozgu
Nedávno sa ukázalo, že magnocelulárne neuróny obsahujúce ACh a lokalizované v bazálnom prednom mozgu (BFM; Basal Forebrain) vyčnievajú monosynapticky do kôry. Učia
Kaudálny-talamo-kortikálny systém
Bazálne gangliá súvisia aj s reguláciou úrovne aktivity organizmu. Ich iný názov je striopallidárny systém, čo je komplex neurónových uzlín, c
Modulačné neuróny
V nervovom systéme bola identifikovaná špeciálna skupina buniek – modulačné neuróny, ktoré samy o sebe nevyvolávajú reakciu, ale regulujú aktivitu iných neurónov. Nadväzujú kontakty s ostatnými
Čo je to pozornosť
Veľké miesto v živote starých Rimanov a Grékov zaujímala rétorika. Ako umenie a ako médium to bolo účinné iba vtedy, ak boli reproduktory dobre umiestnené.
Teórie filtrov
prvý teoretický model pozornosť, alebo model filtra, vytvoril D.E. Broadbent. Predpokladal, že nervový systém napriek mnohým vstupom do určitej miery
Problém pozornosti v tradičnej psychofyziológii
Všeobecnou myšlienkou vyššie opísaných modelov pozornosti je, že na ceste nervových impulzov z receptorov vystavených vonkajším stimulom do kôry,
Problém pozornosti v systémovej psychofyziológii
Tento paradox nevzniká, ak odmietame považovať správanie za reakciu na prezentované podnety. Z hľadiska systémovej psychofyziológie [Shvyrkov, 1995] je správanie realitou
Orientačný reflex
Orientačný reflex alebo reflex "Čo to je?" otvoril I.P. Pavlov. Popísal to ako komplex motorických reakcií, ktoré vznikli pri nečakanom objavení sa nového podnetu.
Približná výskumná činnosť
Orientačnú reakciu (ako naladenie analyzátorov na najlepšie vnímanie nového podnetu) treba odlíšiť od prieskumných reakcií a orientačno-exploračného správania.
Psychofyziológia vedomia
Vedomie je jedným z najzložitejších a zároveň tajomných prejavov mozgovej činnosti. Hoci slovo „vedomie“ je široko používané v každodennej reči
Základné pojmy vedomia
Keď už hovoríme o mozgových základoch psychiky a vedomia, treba povedať, že tieto najzložitejšie prejavy práce mozgu nemožno vysvetliť prácou nejakého samostatného, izolovaného
Re-entry excitácie a syntéza informácií
Vyššie opísaný koncept „svetlého bodu“ pochádza zo skutočnosti, že vedomie je určené určitou úrovňou excitability mozgových štruktúr. Dá sa však predpokladať, že toto
Mozgový základ vnemov
Psychológovia od 20. rokov. je známe, že vnem nastáva skôr neskoro, po 100 ms od momentu prezentácie stimulu (t.j. oveľa neskôr ako príchod zmyslových impulzov do kôry). IN
Mechanizmy myslenia
Náš výskum za posledných desať rokov bol venovaný odpovedi na túto otázku. Úlohou bolo študovať štruktúru kortikálnych spojení pri riešení rôznych mentálnych operácií.
Vedomie, komunikácia a reč
Koncept komunikačnej povahy vedomia prvýkrát predložil P. V. Simonov. Neskôr podobné myšlienky vyjadrili aj iní autori. Oprah
Funkcie vedomia
Otázka funkčného významu subjektívnych skúseností a ich úlohy v správaní je jedným z najdôležitejších problémov vedy o mozgu. Predstavuje výsledok syntézy informácií, mentálnych funkcií
Pojem nevedomia v psychofyziológii
V procese ľudskej činnosti v neustále sa meniacom prostredí sa prichádzajúce informácie spracovávajú na rôznych úrovniach centrálneho nervového systému. Nervózne prepínanie
Indikátory vedomého a nevedomého vnímania
Problém experimentálneho štúdia nevedomého vnímania spočíva v pokusoch identifikovať prahový rozdiel medzi dvoma indikátormi: jedným z nich je indikátor uvedomenia si stimulu; d
Sémantická diferenciácia nevedomých podnetov
Prvýkrát v experimente bol fenomén nevedomého vnímania verbálnych podnetov reprodukovaný skupinou psychológov „New Look“. U zdravých ľudí (študentov) prah op
Dočasné spojenia (asociácie) na nevedomej úrovni
Z pozorovaní psychiatrov je známe, že v určitých prípadoch nevedomé vonkajšie signály, ak sa raz alebo niekoľkokrát zhodovali so silným negatívnym emočným vzrušením
Funkčná asymetria hemisfér a nevedomia
Klasická práca R. Sperryho a jeho kolegov o ľuďoch s „rozštiepeným mozgom“ otvorila cestu neuropsychologickému experimentálnemu štúdiu funkcie
Obráťte časové súvislosti a nevedomie
6.1. Úloha reverzných časových spojení v nervovom mechanizme „psychologickej obrany“ Jedna z foriem psychickej obrany sa prejavuje zvýšením
Význam nevedomých spätnoväzbových stimulov v kognitívnej aktivite
Mnohí výskumníci písali o vplyve nevedomých stimulov na kognitívne funkcie [Kostandov, 1983; Velmans, 1991], hoci neboli vždy opísané jednoznačné výsledky. Tento efekt
Úloha nevedomia v niektorých formách patológie
Vytvorenie podmieneného reflexu na nevedomé vonkajšie podnety vysvetľuje nervový mechanizmus priestorovej orientácie nevidiacich [Beritashvili, 1969]. Základná úloha zvuku
Spánok a sny
1. NÁSTUP AKTÍVNEHO SPÁNKU ALEBO BUDENIE? Už v prvých štúdiách mechanizmov spánku sú jasne načrtnuté dva hlavné body.
Štádiá non-REM spánku a REM spánku
Hlavné údaje získané v priebehu rokov mnohých a rôznorodých štúdií spánku sú nasledovné. Spánok nie je prerušenie činnosti mozgu, je to len iný stav. Na čas
Spánok vo vzťahu a fylogenéze
V ontogenéze sa pomer „spánok a bdelosť“ mení. Takže u novorodencov je stav bdelosti len malá časť dňa a značnú časť spánku zaberá
Potreba spánku
Mnoho ľudí by chcelo spať menej, pretože spánok je podľa nich časom stratený na celý život. Iní by naopak chceli spať viac, pretože sa necítia dostatočne dobre.
spánková deprivácia
Experimenty s depriváciou (umelá spánková deprivácia) naznačujú, že telo potrebuje najmä delta spánok a REM spánok. Po dlhšom nedostatku spánku je hlavná
sny
Sny oddávna udivujú a vzrušujú ľudí. V staroveku boli sny vnímané ako „brány do iného sveta“; verilo sa, že prostredníctvom snov môže dôjsť ku kontaktu s inými svetmi
Dve paradigmy v štúdiu správania a aktivity
So všetkou rozmanitosťou teórií a prístupov používaných v psychológii, psychofyziológii a neurovede ich možno rozdeliť do dvoch skupín. V prvej zo skupín ako hlavný m
Reaktivita
Použitie princípu reaktivity ako vysvetlenie vedecký výskum na základe myšlienok Reného Descarta, ktoré uviedol v prvej polovici 17. storočia. Descartes veril, že organizmus
Aktivita
Uvažovanie o správaní a činnosti ako o činnosti smerujúcej do budúcnosti zahŕňa chápanie činnosti ako základnej vlastnosti živej hmoty; špecifická forma prejavu
Eklekticizmus v psychológii a psychofyziológii
V poslednej dobe sa čoraz viac rozširuje myšlienka aktívnej, cieľavedomej povahy ľudského a zvieracieho správania. Spolu s pozitívnymi dôsledkami tohto
Teória funkčných systémov
2.1. čo je systém? Pojem „systém“ sa zvyčajne používa na označenie zberu, organizácie skupiny prvkov a
Časový paradox
Ako môže výsledok (udalosť, ktorá nastane v budúcnosti) určiť súčasnú aktivitu, byť jej príčinou? Riešením tohto „časového paradoxu“ bol voj
Cieľavedomé správanie
Už u Aristotela bola zrejmá účelovosť správania. Myšlienku účelovosti teda nemožno v žiadnom prípade považovať za novú, hoci v histórii možno rozlíšiť obdobie, kedy
Vedúci odraz
Analýza problémov vzniku a vývoja života z hľadiska TFS priviedla P. K. Anokhina k potrebe zavedenia novej kategórie: anticipačnej reflexie. Vedenie
Teória P.K. Anokhin ako integrálny systém myšlienok
Takže prvá najdôležitejšia výhoda a funkcia, ktorá odlišuje TFS od iných možností systémový prístup, - zavedenie myšlienky výsledku akcie do koncepčnej schémy. Takže asi
Systémové procesy
Ako kľúčové ustanovenia reflexnej teórie P.K. Anokhin vyzdvihol nasledovné: a) výlučnosť počiatočného stimulu ako faktor, ktorý určuje akciu, ktorá je jej
Správanie ako kontinuum výsledkov
Doteraz sme na didaktické účely, ako aj v nadväznosti na tradíciu pôvodnej verzie TFS, používali koncept spúšťacieho podnetu. Je však zrejmé, že využitie tohto pojmu v
Systémové stanovenie aktivity neurónov
3.1. Paradigma reaktivity: neurón, podobne ako jednotlivec, reaguje na podnet
Aktivita ako subjektívna reflexia
Úvaha o vzťahu medzi jednotlivcom a prostredím z hľadiska TFS už dlho viedla k záveru, že behaviorálne kontinuum je úplne obsadené procesmi organizácie a implementácie funkcie.
Fyzické vlastnosti prostredia a cielené správanie
Pred viac ako 30 rokmi J. Lettvin a kol., ktorí študovali vzťah medzi aktivitou žabích neurónov sietnice a jej správaním, sformulovali vo veľmi živej forme svoju predstavu o tom, čo
Závislosť aktivity centrálnych a periférnych neurónov od cieľa správania
Presvedčivé príklady toho, ako sa subjektivita reflexie prejavuje v organizácii mozgovej aktivity, možno získať analýzou závislosti aktivity neurónov od cieľov správania.
Význam eferentných vplyvov
Súvislosť medzi aktivitou gangliových buniek sietnice a správaním so zavretými očami je daná už spomínanými eferentnými vplyvmi. Už začiatkom tohto storočia S. Ramon y Cajal
Psychofyziologický problém a úlohy systémovej psychofyziológie
V tejto časti odpovieme na nasledujúce otázky. Ako závisia úlohy psychofyziológie od metodických pokynov? Je medzi nimi nejaká špecifická úloha psycho
Korelatívna psychofyziológia
Tradičné psychofyziologické štúdie sa spravidla vykonávajú z hľadiska „korelatívnej (porovnávacej) psychofyziológie“. V týchto štúdiách mentálne
Systémové riešenie psychofyziologického problému
Podstata systémového riešenia psychofyziologického problému spočíva v nasledujúcom ustanovení. Duševné procesy charakterizujúce organizmus a správanie pôsobia ako celok a neurofyziológovia
Úlohy systémovej psychofyziológie a jej význam pre psychológiu
Použitie vyššie uvedeného riešenia psychofyziologického problému v systémovej psychofyziológii ako jednej z najdôležitejších zložiek metodológie umožňuje vyhnúť sa redukcionizmu.
Interakcia korelačnej a systémovej psychofyziológie
Filozofia vedy potvrdzuje užitočnosť koexistencie alternatívnych teórií, čo prispieva k ich vzájomnej kritike a urýchľuje rozvoj vedy. Dobrý príklad férovosti
Systemogenéza
V predchádzajúcom odseku pri formulovaní úloh systémovej psychofyziológie nie je náhodou na prvom mieste úloha študovať formovanie systémov. Ďalej uvidíme, že história
Organogenéza a systemogenéza
Na rozdiel od koncepcie organogenézy, ktorá predpokladá postupný vývoj jednotlivých morfologických orgánov, ktoré plnia zodpovedajúce lokálne „súkromné“ funkcie, koncepcia systému
Učenie ako reaktivácia vývinových procesov
V súčasnosti sa všeobecne uznáva, že mnohé vzory modifikácií funkčných a morfologické vlastnosti neurónmi, ako aj regulácia génovej expresie, ktoré spočívajú v o
Systémová špecializácia a systémová špecifickosť neurónov
Špecializácia neurónov vo vzťahu k novovzniknutým systémom - systémová špecializácia - je konštantná, t.j. neurón je systémovo špecifický. V súčasnosti
Historické určenie úrovne organizácie systémov
Predstavy o vzorcoch rozvoja rozvíjajú mnohí autori v súvislosti s myšlienkami organizácie úrovní (pozri [Anokhin, 1975, 1980; Rogovin, 1977; Aleksandrov, 1989, 1995,
Štruktúra subjektívneho sveta a subjekt správania
Špecializácia neurónov vo vzťahu k prvkom individuálnej skúsenosti znamená, že ich činnosť odráža nie vonkajší svet ako taký, ale vzťah jednotlivca k nemu (pozri aj pár
Dynamika subjektívneho sveta ako zmena stavov subjektu správania
Z týchto pozícií možno dynamiku subjektívneho sveta charakterizovať ako zmenu stavov subjektu správania v priebehu nasadenia behaviorálneho kontinua (pozri obr.
Modifikovateľnosť systémovej organizácie behaviorálneho aktu v následných implementáciách
Dokonca aj F. Bartlett navrhol úplne zavrhnúť názory, podľa ktorých sa „reprodukcia z pamäte“ považuje za „opätovné vybudenie nezmenených“ stôp“, objavenie sa v repertoári jednotlivca nového
Fixovanie etáp učenia vo forme prvkov zážitku
Po načrtnutí základného prístupu k výberu prvkov subjektívnej skúsenosti vytvorila teória funkčných systémov základ experimentov na štúdium subjektívnej fragmentácie správania a
Vplyv histórie učenia na štruktúru skúseností a organizáciu mozgovej činnosti
Subjektívne kontinuum, podobne ako kontinuum správania, je lineárna postupnosť po sebe nasledujúcich stavov, ktoré zodpovedajú aktom správania. Tieto posuny
Mozgové potenciály súvisiace s udalosťami
ERP sú širokou triedou elektrofyziologických javov, ktoré sa izolujú od „pozadia“ alebo „surového“ elektroencefalogramu (EEG) špeciálnymi metódami. Termín SSP
Stručná história metódy SSP
Spojenie elektrickej aktivity mozgu s dianím v prostredí a správaním prvýkrát predviedol a opísal Angličan R. Caton (Richard Caton) v rokoch 1875–1887. a n
Všeobecná charakteristika signálu
SSP sa izoluje pomocou špeciálnych metód z EEG. Frekvenčný rozsah SSP zahŕňa pásmo od 0 Hz do 3 kHz a je obmedzený na jednej strane ultrapomalou elektrickou aktivitou
Štandardné metódy na získanie reprodukovateľnej konfigurácie SSP
Metodické požiadavky na EEG záznam (inštalácia elektród, výber záznamového systému, šírky pásma zosilňovačov, metódy eliminácie artefaktov) sú popísané v kapitole 2. Upozorňujeme, že h
Priemerovanie
Nasledujúce predpoklady sú základom výberu ERP zo signálu EEG: a) v situácii opakovaného opakovania udalosti zaznamenaný signál EEG (SUMi (t))
Filtrácia
Náhodný, "šum" komponent jedinej implementácie ERP ("surové" EEG) možno eliminovať vyhladením. ALGEBRAICKÉ FILTRE
Opis SSP
V dôsledku akumulácie segmentov EEG spojených s určitými udalosťami, ich spriemerovaním, digitálnym filtrovaním alebo inými postupmi sa získa SSP - krivka, ktorá je opísaná ako po
Vlastnosti metódy SSP
V posledných desaťročiach boli vyvinuté metódy na zaznamenávanie mozgovej aktivity, ktoré majú významný výskumný potenciál (pozri tiež kapitolu 2). Avšak pri vývoji nových
Vizuálne evokované potenciály
Vizuálne evokované potenciály (VEP, visual evoked potentials - VEP) [Shagas, 1975; Rutman, 1979; Maksimová, 1982; Rockstroh et al., 1982] sú registrovaní v situácii vizuálnej prezentácie
sluchové evokované potenciály
Sluchové evokované potenciály (AEP) [Shagas, 1975; Rutman, 1979; Rockstroh a kol., 1982; Hughes, 1985] sú registrovaní v situácii prezentácie fám
Somatosenzorické evokované potenciály
Somatosenzorické evokované potenciály (SEP) [Shagas, 1975; Rutman, 1979; Rockstroh et al., 1982] sú registrovaní v situácii srsti
Potenciály spojené s vykonávaním pohybov
Potenciály spojené s výkonom pohybov (PSVD, pohybové potenciály - MRP, pohybové mozgové potenciály - MRBP; v ruskojazyčnej literatúre sa používa
Podmienená negatívna vlna
Podmienená negatívna vlna (UNV, podmienená negatívna variácia - CNV alebo očakávaná vlna - E-vlna). V situácii prezentácie dvoch podnetov, prvý z
Princípy zefektívnenia fenomenológie SSP
Zoznam známych typov SSP sa neustále aktualizuje a nie je dôvod považovať ho za takmer dokončený. Uveďme ako príklad najznámejší fenomén
Problém funkčného významu csp
V rámci korelačnej psychofyziológie (pozri [Shvyrkov, 1995] a kap. 14) sa predpokladá, že fluktuácie (komponenty) ERP odrážajú špecifické funkcie mozgových štruktúr, ktoré implementujú
Psychologické koreláty
Hľadanie psychologických korelátov SSP ukázalo, že: 1) rovnaký SSP je spojený s mnohými psychologickými procesmi (funkciami) a 2) rovnakým mentálnym fu
Hľadanie mozgových zdrojov ERP ukázalo nasledovné: 1) akékoľvek kolísanie ERP zaznamenané z povrchu hlavy je odrazom aktivity mnohých kortikálnych a subkortikálnych
CSP ako odraz dynamiky individuálnej skúsenosti
SSP sú celkový elektrický potenciál rôznych zložiek mozgového tkaniva, ku ktorému prispievajú neuróny (soma, dendrity a axóny), gliové bunky, bunkové membrány
Potenciál pre všestrannú konfiguráciu
Porovnanie SSP sprevádzajúceho správanie subjektov v rôznych experimentálnych situáciách ukazuje, že implementácia a zmena behaviorálneho aktu zodpovedá potenciálu univerzity.
Perspektívy využitia ssp
Efektívnosť použitia SSP ako metódy psychofyziologického výskumu je daná riešením hlavného problému: aký je pomer parametrov SSP, mozgovej aktivity, f.
Pojem vlastností nervového systému
Problém individuálnych psychologických rozdielov medzi ľuďmi bol v ruskej psychológii vždy považovaný za jeden zo základných. Najväčší príspevok k rozvoju
Všeobecné vlastnosti nervovej sústavy a integrálna formálno-dynamická charakteristika individuality
Pre experimentálne overenie rozvinutých predstáv o vlastnostiach nervového systému a ich psychologických prejavoch V.D. Nebylitsyn uskutočnil štúdiu fyziologických základov inteligencie
Integrálna individualita a jej štruktúra
Prístup vyvinutý B. C. Merlinom vytvoril základ pre rozvoj pôvodnej školy štúdia povahy temperamentu. Predpoklady, na ktorých bol tento prístup založený
Individuálne charakteristiky správania e zvierat
Dôležitým trendom, ktorý sa objavil vo vývoji problému charakteru individuálnych psychologických rozdielov po B.M. Teplovej a V.D. Nebylitsyn a bol založený na modeli správania
Integrácia poznatkov o osobnosti
Koncom 80. rokov. s cieľom vytvoriť novú stratégiu skúmania povahy individuálnych psychologických rozdielov medzi ľuďmi. M. Rusalov vypracoval metodiku hodnotenia typu dotazníka
Medzikultúrne štúdie osobnosti
IN posledné roky v diferenciálnej psychofyziológii sa začala uplatňovať metodológia medzikultúrneho výskumu. Medzikultúrny diferenciálny psychofyziologický výskum
Psychofyziológia profesionálnej činnosti
Na hraniciach prírodných vied a psychológie sa sformovalo množstvo špeciálnych vedných disciplín a smerov, medzi ktoré patrí psychológia práce, inžinierska psychológia a ergonómia, ktoré sú predmetom tzv.
Teoretické základy využitia psychofyziológie pri riešení praktických problémov psychológie práce
Pochopenie potreby a vyhliadok na štúdium psychofyziologických procesov v profesionálnej činnosti je uľahčené myšlienkami, ktoré berú do úvahy mentálne a fyziológovia.
Metodologická podpora psychofyziologického aspektu aplikovaného výskumu
V aplikovanom výskume je komplexnosť psychofyziologického výskumu zabezpečená využitím metódy polyefektora, ktorá zahŕňa registráciu frekvencie
Psychofyziológia profesionálneho výberu a profesionálnej vhodnosti
Použitie psychofyziologických metód v psychológii práce bolo spôsobené potrebou vyvinúť objektívne a kvantitatívne kritériá pre psychofyziologický výber, ktoré ako
Psychofyziologické zložky pracovnej kapacity
Schopnosť osoby vykonávať určitú činnosť v daných časových limitoch a výkonnostných parametroch určuje obsah pracovnej schopnosti ako hlavnej zložky.
Psychofyziologické determinanty adaptácie človeka na extrémne podmienky činnosti
V súčasnosti sa hlavnými smermi v štúdiu adaptácie stali definície štádií formovania psychofyziologického systému adaptácie, kritérií pre jeho formovanie, alokácie.
Psychofyziologické funkčné stavy (PFS)
Relevantnosť štúdia PFS je určená ich prínosom k zabezpečeniu výkonnosti a spoľahlivosti človeka, ako aj nárastom počtu profesií a zmenami pracovných podmienok.
biofeedback (bos)
záujem o biologický výskum spätná väzba(BOS) za účelom svojvoľnej kontroly FS na základe objektívnych informácií o dynamike psychofyziologických
Psychofyziologický rozbor obsahu odbornej činnosti
Psychofyziologická analýza profesionálnej činnosti zahŕňa „považovanie za komplexný, viacrozmerný a viacúrovňový, dynamický a rozvíjajúci sa fenomén“ [Lomov,
Porovnávacia psychofyziológia
Porovnávacia psychofyziológia je veda zameraná na stanovenie vzorcov a identifikáciu rozdielov v štrukturálnych a funkčná organizácia mozog, správanie a psychika zvierat
Príchod psychiky
V súčasnosti je najviac akceptovaný názor, že psychika je atribútom živých systémov a neživá príroda, vrátane zložitých zariadení vytvorených ľuďmi, nie je o psychike.
Evolúcia druhov
Moderné údaje o vývoji živočíchov naznačujú divergenciu evolučných línií a vývoj paralelných línií (obr. 19.2 A), a to aj medzi cicavcami (obr. 19.2 B) a medzi
Evolučné premeny mozgu
Štruktúra mozgu u zvierat odlišné typy pestrá. A hoci, ako vyplýva z obr. 19.3, príbuzné druhy, napríklad medzi kôrovcami alebo cicavcami, majú spoločné znaky v štruktúre
Porovnávacia metóda v systémovej psychofyziológii
Systémová psychofyziológia, ktorej základy položili práce V.B. Shvyrkov a jeho kolegovia, je založená na rozpoznaní: 1) jedinej psychofyziologickej reality, v ktorej psychologické
FYZIOLÓGIA ROVNOVÁHY, SLUCHU A REČI
R. Klinke
Táto kapitola je venovaná fyziológii dvoch fylogeneticky príbuzných zmyslových orgánov, sluchu a rovnováhu. Sú nielen anatomicky úzko spojené, nachádzajú sa vedľa seba v skalnej kosti a tvoria sa vnútorné ucho, ale vyskytli sa aj v priebehu evolúcie z jednej štruktúry. Keďže najdôležitejší dorozumievací prostriedok pre človeka – reč – sprostredkúva orgán sluchu, fyziológia reči diskutované aj v tejto kapitole.
Reč si vyžaduje sluch. Okrem toho je verbálna komunikácia najdôležitejším prostriedkom učenia, preto je hluchota či dokonca nedoslýchavosť najvážnejšou hrozbou pre duševný vývoj dieťaťa. Porovnávacie fyziologické štúdie ukázali, že hluchota ho postihuje viac ako slepota. Preto je sluch pre človeka najdôležitejším zmyslom.
Fyziológia periférneho senzorického aparátu
Úvodné anatomické komentáre . Jednou zo zložiek je vestibulárny orgán membránový labyrint, generovanie vnútorné ucho; jeho ďalšou zložkou je orgán sluchu (obr. 12.1). Membranózny labyrint je naplnený tekutinou endolymfa, a ponorený do iného, tzv perilymfa. Vestibulárny orgán pozostáva z dvoch morfologických podjednotiek - otolitu prístroja ( macula utriculi a macula sacculi ) A polkruhové kanály (vpredu A zadné vertikálne A horizontálne kanály). V oblasti makúl (škvrny) a v polkruhových kanálikoch v blízkosti ampuliek sa nachádza senzorický epitel obsahujúci receptory, ktorý je pokrytý rôsolovitou hmotou vytvorenou najmä mukopolysacharidy. V otolitickom aparáte táto hmota pokrýva zmyslové bunky ako vankúš a obsahuje usadeniny uhličitanu vápenatého vo forme drobných kryštály kalcitu(otolity). Vďaka prítomnosti týchto "kamenných" inklúzií sa nazýva otolitická membrána. Doslovný preklad gréckeho výrazu " otolit hus "-" ucho kameň“. V polkruhových kanálikoch je rôsolovitá hmota skôr ako membránová priehradka. Táto štruktúra kopula, neobsahuje kryštály.
Receptory a adekvátny stimul . V senzorickom epiteli makuly a polkruhových kanálikov sa nachádzajú dva morfologicky odlišné typy receptorových buniek, ktoré sa zjavne výrazne nelíšia vo svojich fyziologických vlastnostiach.
Oba typy buniek nesú na svojom voľnom povrchu submikroskopické chĺpky. (cilia), preto sa nazývajú chlpaté (obr. 12.2). Pomocou elektrónového mikroskopu je možné rozlišovať stereocília(60-80 na každej receptorovej bunke) a kinocília(jeden za druhým). Receptory sú sekundárne senzorické bunky, t.j. nenesú vlastné nervové procesy, ale sú inervované aferentnými vláknami neurónov. vestibulárny ganglion, tvoriaci vestibulárny nerv. Eferentné vlákna tiež končia na receptorových bunkách. Aferentné prenášajú informácie o úrovni excitácie receptorov do centrálneho nervového systému a eferenty menia ich citlivosť, ale význam tohto vplyvu stále nie je celkom jasný. Ukázala ich registrácia aktivity jednotlivých aferentných vlákien vestibulárneho nervu
Ryža. 12.1.Schéma vestibulárneho labyrintu. Jeho lymfatické priestory komunikujú s kochleárnym

Ryža. 12.2.Schéma dvoch receptorových buniek zmyslového epitelu vestibulárneho orgánu a ich nervových vlákien. Keď je zväzok riasiniek naklonený smerom ku kinocíliu, frekvencia impulzov v aferentnom nervovom vlákne sa zvyšuje a pri naklonení v opačnom smere sa znižuje.
pomerne vysoká pravidelná oddychová aktivita, tie. impulzov aj pri absencii vonkajších podnetov. Ak je rôsolovitá hmota experimentálne posunutá voči senzorickému epitelu, takáto aktivita sa zvyšuje alebo znižuje v závislosti od smeru posunu. Tieto zmeny prebiehajú nasledujúcim spôsobom. Keďže riasinky sú ponorené do rôsolovitej hmoty, pri pohybe sa odchyľujú. Posun ich zväzku slúži ako adekvátny stimul pre receptor. Keď smeruje ku kinocíliu (obr. 12.2), aktivuje sa príslušné aferentné vlákno: rýchlosť jeho impulzu sa zvyšuje. Pri posunutí v opačnom smere sa frekvencia impulzov znižuje. Posun v smere kolmom na túto os nemení aktivitu. Informácie sa prenášajú z receptorovej bunky do zakončenia aferentného nervu vďaka receptorovému potenciálu a zatiaľ neidentifikovanému neurotransmiteru. Najdôležitejšia vec je tu posun(prehnutie) mihalnice je adekvátnym stimulom pre vestibulárne receptory, čím sa zvyšuje alebo znižuje (v závislosti od jej smeru) aktivita aferentného nervu. Existuje teda morfologická (podľa umiestnenia mihalníc) a funkčná
(podľa charakteru vplyvu na činnosť) orientácia receptorová bunka.
Prirodzené podnety pre makulu . Ako už bolo uvedené, riasinky receptorových buniek sú ponorené do otolitickej membrány. V druhom prípade je v dôsledku prítomnosti kryštálov kalcitu hustota (približne 2,2) výrazne vyššia ako hustota endolymfy (asi 1), ktorá vypĺňa zvyšok vnútornej dutiny sacculus (guľovitý vak) a utriculus (eliptický vak, maternica). Preto v dôsledku všadeprítomného gravitačného zrýchlenia, kedykoľvek senzorický epitel otolitového aparátu nezaujme dokonale horizontálnu polohu, gravitačná sila spôsobuje posúvanie (na veľmi malú vzdialenosť) pozdĺž nej celej otolitickej membrány. (Predstavte si, čo by sa stalo, keby rôsolovitá hmota, znázornená červenou farbou na obrázku 12.2, bola veľmi ťažká a učebnicu by ste držali zvisle a naklonili ju nabok. Prirodzene, že by sa kĺzala pod uhlom.) Tento pohyb sa ohýba riasinky, tj. na receptory pôsobí adekvátny stimul. Keď človek stojí vzpriamene a jeho hlava je v „normálnej“ polohe, makula utrikulu je takmer horizontálna a otolitická membrána nepôsobí šmykovou silou na ňou pokrytý zmyslový epitel. Keď je hlava naklonená, makula utriculus je v uhle k horizontu, jej riasy sú ohnuté a receptory sú stimulované. V závislosti od smeru sklonu sa frekvencia impulzov eferentného nervu buď zvyšuje alebo znižuje. Situácia s makulou sacculus je v princípe podobná, ale v normálnej polohe hlavy je umiestnená takmer vertikálne (obr. 12.1). Pri akejkoľvek orientácii lebky teda každá z otolitových membrán svojím spôsobom ovplyvňuje zmyslový epitel a vzniká špecifický vzor excitácie nervových vlákien. Keďže v každej makule sú dve populácie receptorových buniek s opačne orientovanými riasinkami, nemožno povedať, že naklonenie hlavy týmto smerom aktivuje aferentáciu. Naopak, v každom prípade sú niektoré vlákna aktivované, zatiaľ čo iné sú inhibované. Takáto poloha hlavy, pri ktorej by aktivita všetkých nervových vlákien klesla na nulu, neexistuje.
Centrálne zložky vestibulárneho systému, hodnotiace typ excitácie vestibulárneho nervu, informujú telo o orientácii lebky v priestore. Poskytovanie takýchto informácií je najdôležitejšou funkciou otolitových orgánov. Gravitačné zrýchlenie je len jednou špeciálnou formou lineárnych zrýchlení; Prirodzene, makuly reagujú aj na ostatných. Gravitačné zrýchlenie je však také veľké, že v jeho prítomnosti iné

Ryža. 12.3.Schéma ľavého horizontálneho polkruhového kanála (pohľad zhora). S výnimkou vydutiny predstavujúcej utriculus nie sú ostatné časti labyrintu zobrazené. Uhlové zrýchlenie v naznačenom smere čierna šípka(predstavte si, že učebnicu otáčate týmto smerom), vychýli kupolu po ceste červená šípka
lineárne zrýchlenia vyskytujúce sa v Každodenný život(napríklad pri akcelerácii auta), zohrávajú podriadenú úlohu pre vestibulárny systém a môžu byť dokonca nesprávne interpretované centrálnym nervovým systémom.
Prirodzené podnety pre polkruhové kanáliky . Druhý typ adekvátnych stimulov pre mihalnice vestibulárnych receptorov vnímame v polkruhových kanálikoch (obr. 12.3). Hoci ich skutočný tvar v tele nie je dokonalý kruh (obr. 12.1), pôsobia ako uzavreté kruhové trubice naplnené endolymfou. V oblasti ampulky je ich vonkajšia stena vystlaná senzorickým epitelom (obr. 12.3); tu do endolymfy vyčnieva kupula s riasinkami receptorových buniek hlboko v nej uložených. Kupula bez minerálov polkruhových kanálikov má presne rovnakú hustotu ako endolymfa. V dôsledku toho lineárne zrýchlenie (vrátane gravitačného) neovplyvňuje tento orgán; pri priamočiarom pohybe a rôznych orientáciách hlavy zostávajú vzájomné polohy polkruhových kanálikov, mihalnice a mihalníc nezmenené. Iný vplyv na rohu(otočný) zrýchlenie. Pri otáčaní hlavičky sa s ňou prirodzene otáčajú aj polkruhové kanáliky, no endolymfa svojou zotrvačnosťou zostáva v prvom momente na svojom mieste. Na oboch stranách kuply, spojenej so stenou kanála a tvoriacej vodotesnú bariéru, vzniká tlakový rozdiel, v dôsledku čoho sa odchyľuje v smere proti pohybu (obr. 12.3). To spôsobí, že na mihalnice pôsobí šmyková sila a tým sa mení činnosť aferentného nervu. V horizontálnych kanáloch sú všetky receptory orientované tak, že kinocília smeruje k utriculus, takže aferentná aktivita sa zvyšuje, keď sa kupula odchyľuje rovnakým smerom. (utrikulopetálne). V ľavom horizontálnom polkruhovom kanáli k tomu dochádza pri otáčaní doľava. Vo vertikálnych kanáloch sa aferentácia aktivuje, keď utriculo-fugal odchýlka kopule (od utriculus). Impulz všetkých týchto vlákien, prichádzajúci z troch kanálov na každej strane, je tiež odhadnutý CNS a poskytuje informácie o uhlových zrýchleniach pôsobiacich na hlavu. Práve preto, že hlava sa môže otáčať okolo troch priestorových osí – nakláňať dopredu a dozadu, doľava a doprava a otáčať sa okolo dlhej osi tela – sú to práve tri polkruhové kanáliky, ktoré ležia v troch rovinách takmer na seba kolmých. Pri otáčaní okolo akejkoľvek diagonálnej osi je stimulovaných viac ako jeden kanál. Zároveň mozog vykonáva vektorovú analýzu informácií a určuje skutočnú os rotácie. V klinických štúdiách je dôležité vziať do úvahy, že takzvaný horizontálny polkruhový kanál nie je úplne horizontálny: jeho predný okraj je vyvýšený približne o 30°.
Vlastnosti kupulovej mechaniky. Uvažujme najskôr, čo sa deje s kupolou pri krátkodobom uhlovom zrýchlení, t.j. keď len otočíme hlavy. Ako vyplýva z obr. 12.4, A, vychýlenie kopule nezodpovedá tomuto zrýchleniu, ale okamžitej uhlovej rýchlosti. Zmeny vo frekvencii neurónových impulzov v porovnaní so spontánnymi sa teda približujú skôr zmenám uhlovej rýchlosti než uhlovému zrýchleniu, hoci sily, ktoré spôsobujú deformáciu kuply, sú spôsobené práve zrýchlením. Po dokončení tohto krátkeho pohybu sa kupula vráti do pôvodného stavu a činnosť aferentného nervu sa zníži na pokojovú úroveň. Na obr. 12.4, B ukazuje sa zásadne iná situácia, ktorá sa pozoruje pri dlhodobej rotácii (napríklad v centrifúge), keď sa po počiatočnom zrýchlení na dlhú dobu ustáli konštantná uhlová rýchlosť. Kupula, v prvom momente vychýlená, sa potom pomaly vracia do pokojovej polohy. Rýchle zastavenie rovnomernej rotácie ju opäť vychýli, ale v opačnom smere (v dôsledku zotrvačnosti sa endolymfa ďalej pohybuje, čo vedie k tlakovému rozdielu na oboch stranách kuply, čo vedie k jej posunutiu, ktorého charakteristika s s výnimkou smerovosti, sú rovnaké ako na začiatku pohybu). Trvá pomerne dlhý čas (10–30 s), kým sa kupla vráti do pôvodnej polohy.

Ryža. 12.4.Odchýlka kopule a aktivita aferentného nervového vlákna: A - s krátkym otočením (napríklad hlavy); B-s predĺženým otáčaním (napríklad na stoličke). Všimnite si rozdiel v časovej škále na obrázkoch
Rozdiel medzi odozvami kuply na krátkodobú a dlhodobú stimuláciu súvisí s mechanickými vlastnosťami systému kupla-endolymfa, ktorý sa v prvej aproximácii správa ako silne tlmené torzné kyvadlo. Zároveň by sa malo pamätať na to, že sily, ktoré vychyľujú kupolu, vždy sú spôsobené zrýchlením, aj keď pri krátkodobých uhlových zrýchleniach, najbežnejších za fyziologických podmienok, je jeho odchýlka úmerná nie im, ale uhlovej rýchlosti.
Deformácie kopule sú zvyčajne veľmi malé, ale jej receptory sú mimoriadne citlivé. Pri pokusoch na zvieratách sa pre nich ukázala ako nadprahová stimulácia veľmi rýchla rotácia tela len o 0,005° (vychýlenie kuply - rovnakého rádu).
Primárne aferenty vestibulárneho nervu končia hlavne v oblasti vestibulárnych jadier. medulla oblongata. Na každej strane tela sú štyri, líšia sa od seba anatomicky aj funkčne: horný(Bekhterev), mediálne(Schwalbe), bočné(Deiters) a dno(Valec). Impulzy prichádzajúce k nim z vestibulárnych receptorov samy o sebe neposkytujú presné informácie o polohe tela v priestore, pretože uhol natočenia hlavy v dôsledku pohyblivosti krčných kĺbov nezávisí od orientácie tela. . Centrálny nervový systém musí brať do úvahy aj polohu hlavy voči telu. V dôsledku toho dostávajú vestibulárne jadrá dodatočnú aferentáciu z cervikálne receptory(svaly a kĺby). Pri experimentálnej blokáde týchto spojení dochádza k rovnakým nerovnováham ako pri poškodení labyrintu. Vestibulárne jadrá prijímajú somatosenzorické signály aj z iných kĺbov (nohy, ruky).
Nervové vlákna vychádzajúce z týchto jadier sú spojené s inými časťami centrálneho nervového systému, ktorý poskytuje reflexy na udržanie rovnováhy. Tieto cesty zahŕňajú nasledujúce.
ale. vestibulospinálny trakt, vlákna ktorých v konečnom dôsledku ovplyvňujú hlavne γ motorické neuróny extenzorových svalov, hoci tiež končia v α motorických neurónoch.
b. Odkazy s motoneuróny krčnej miechy, v zásade súvisí s vestibulospinálnym traktom.
v. Odkazy s okulomotorické jadrá, ktoré sprostredkúvajú pohyby očí spôsobené vestibulárnou činnosťou. Tieto vlákna prebiehajú ako súčasť mediálneho pozdĺžneho zväzku.
d) Cesty smerujúce do vestibulárne jadrá na opačnej strane mozgu, umožňujúce spoločné spracovanie aferentácie z oboch strán tela.
e) Prepojenie s cerebellum najmä s archcerebellum (pozri nižšie).
e) Prepojenie s retikulárna formácia, zabezpečenie dopadu na retikulospinálny trakt - ďalšia (polysynaptická) cesta k α- a γ-motorickým neurónom.
dobre. Cesty prechádzajúce cez thalamus v postcentrálny gyrus mozgová kôra, ktorá vám umožňuje spracovávať vestibulárne informácie, a teda vedome sa pohybovať v priestore.
h. Vlákna smerujú k hypotalamus, podieľajú sa najmä na výskyte kinetózy. Tento súbor spojení, iba tie hlavné, ktoré sú uvedené vyššie, umožňuje vestibulárnemu systému hrať ústrednú úlohu pri vytváraní motorickej eferencie, ktorá zabezpečuje udržanie požadovanej polohy tela a zodpovedajúcich okulomotorických reakcií. V čom vzpriamený postoj a chôdza determinovaný hlavne otolitickým aparátom, kým polkruhové kanáliky riadia hlavne smer pohľadu. Práve aferentácia z polkruhových kanálikov spolu s okohybnými mechanizmami zabezpečuje vizuálny kontakt s okolím pri pohyboch hlavy. Pri jej otočení alebo naklonení sa oči pohybujú opačným smerom, takže obraz na sietnici sa nemení (pozri statokinetické reflexy). Horizontálne kompenzačné pohyby očí sú riadené horizontálnym polkruhovým kanálom, vertikálne predným vertikálnym kanálom a ich rotácia je hlavne zadným vertikálnym kanálom.
Ďalšou dôležitou súčasťou CNS, ktorá sa podieľa na týchto procesoch, je mozoček, ktorý okrem sekundárnych diskutovaných vyššie prijíma niektoré primárne vestibulárne aferenty (tzv. priama senzorická cerebelárna dráha). Všetky u cicavcov v nej končia machovými vláknami v bunkách - zrnkami uzliny ( nodulus) a skartovať (flokulus ) súvisiaci so starovekým mozočkom ( archicerebellum ), a čiastočne uvula ( uvula) a okolochka (paraflokulus ) starý mozoček ( paleocerebellum ). Granulové bunky majú excitačný účinok na Purkyňove bunky v rovnakých oblastiach a ich axóny smerujú opäť do vestibulárnych jadier. Takýto obvod vykonáva jemné „ladenie“ vestibulárnych reflexov. Pri cerebelárnej dysfunkcii sú tieto reflexy dezinhibované, čo sa prejavuje napríklad zvýšeným alebo spontánnym nystagmom (pozri nižšie), nerovnováhou, prejavujúcou sa sklonom k pádu, neistou chôdzou a nadmerným rozsahom pohybu najmä pri chôdzi („kohútikový krok "). Uvedené príznaky súvisia so syndrómom cerebelárna ataxia.
Typy neurónových impulzov vestibulárnych jadier sú také rozmanité ako ich kontakty, takže ich podrobne nezvažujeme. Podrobnosti možno nájsť v odbornej literatúre.
vestibulárne reflexy; klinické testy
Statické a statokinetické reflexy . Rovnováha je udržiavaná reflexne, bez zásadnej účasti vedomia na tomto. Prideliť statické a statokinetické reflexy. S oboma sú spojené vestibulárne receptory a somatosenzorická aferentácia, najmä z cervikálnych proprioceptorov. Statické reflexy zabezpečujú primeranú vzájomnú polohu končatín, ako aj stabilnú orientáciu tela v priestore, t.j. posturálne reflexy. Vestibulárna aferentácia pochádza v tomto prípade z otolitických orgánov. Statický reflex, ľahko pozorovateľný u mačky vďaka svojmu vertikálnemu tvaru zrenice, kompenzačná rotácia očnej gule pri otáčaní hlavy okolo dlhej osi tela (napríklad ľavé ucho dole). Žiaci zároveň po celý čas zachovávajú polohu veľmi blízko vertikále. Tento reflex sa pozoruje aj u ľudí. Statokinetické reflexy sú reakcie na motorické podnety, ktoré sa samy prejavujú v pohyboch. Sú spôsobené excitáciou receptorov polkruhových kanálikov a otolitických orgánov; Príkladmi sú rotácia mačacieho tela pri páde, ktorá zabezpečí, aby dopadla na všetky štyri nohy, alebo pohyb človeka, ktorý znovu nadobudne rovnováhu po tom, čo zakopne.
Jeden zo statokinetických reflexov je vestibulárny nystagmus– podrobnejšie sa budeme zaoberať v súvislosti s jeho klinickým významom. Ako je uvedené vyššie, vestibulárny systém spôsobuje rôzne pohyby očí; nystagmus, ako ich špeciálna forma, sa pozoruje na začiatku rotácie, ktorá je intenzívnejšia ako bežné krátke otáčky hlavy. Ako sa oči otáčajú proti smery otáčania, aby zachovali pôvodný obraz na sietnici, pred dosiahnutím jej krajnej možnej polohy však prudko "poskočia" v smere otáčania a v zornom poli sa objaví ďalší úsek priestoru. Potom ich nasleduje pomaly spätný pohyb.
Pomalú fázu nystagmu spúšťa vestibulárny systém a rýchle „skákanie“ pohľadu spúšťa prepontínna časť retikulárnej formácie.
Pri rotácii telesa okolo zvislej osi dochádza k podráždeniu takmer len horizontálnych polkruhových kanálikov, t.j. vychýlenie ich kupuliek spôsobuje horizontálny nystagmus. Smer oboch jeho zložiek (rýchla a pomalá) závisí od smeru rotácie a tým aj od smeru deformácie kupule. Ak sa telo otáča okolo horizontálnej osi (napríklad pri prechode cez uši alebo sagitálne cez čelo), dochádza k stimulácii zvislých polkruhových kanálikov a vzniku vertikálneho alebo rotačného nystagmu. Smer nystagmu je zvyčajne určený jeho rýchla fáza, tie. pri „pravom nystagme“ pohľad „skočí“ doprava.
Pri pasívnej rotácii tela vedú k výskytu nystagmu dva faktory: stimulácia vestibulárneho aparátu a pohyb zorného poľa vzhľadom na osobu. Optokinetický (spôsobený zrakovou aferentáciou) a vestibulárny nystagmus pôsobia synergicky. Neurónové spojenia s tým spojené sú diskutované vyššie.
Diagnostická hodnota nystagmu . Nystagmus (bežne označovaný ako „postrotačný“) sa klinicky používa na testovanie vestibulárnych funkcií. Subjekt sedí v špeciálnom kresle, ktoré sa dlho otáča konštantnou rýchlosťou a potom sa náhle zastaví. Na obr. 12.4 ukazuje správanie kopule. Zarážka spôsobí, že sa vychýli v opačnom smere, než v akom sa odchýlil na začiatku pohybu; výsledkom je nystagmus. Jeho smer môže byť určený registráciou deformácie kopule; to by malo byť opak smer predchádzajúceho pohybu. Záznam očných pohybov sa podobá záznamu získanému v prípade optokinetického nystagmu (pozri obr. 11.2). To sa nazýva nystagmogram.
Po testovaní na postrotačný nystagmus je dôležité túto možnosť vylúčiť fixácia pohľadu v jednom bode, keďže pri okulomotorických reakciách dominuje zraková aferentácia nad vestibulárnou a za určitých podmienok je schopná potlačiť nystagmus. Preto je predmet kladený na Frenzelove okuliare s vysoko konvexnými šošovkami a vstavaným zdrojom svetla. Robia ho „krátkozrakým“ a neschopným fixovať svoj pohľad, pričom lekárovi umožňujú ľahko pozorovať pohyby očí. Takéto okuliare sa vyžadujú aj pri teste na prítomnosť spontánna nystagmus - prvý, najjednoduchší a najviac dôležitý postup v klinickej štúdii vestibulárnej funkcie.
Ďalším klinickým spôsobom, ako spustiť vestibulárny nystagmus, je tepelná stimulácia horizontálne polkruhové kanály. Jeho výhodou je možnosť otestovať každú stranu tela zvlášť. Hlava sediaceho je naklonená dozadu približne o 60° (u ležiaceho na chrbte je zdvihnutá o 30°), takže horizontálny polkruhový kanál je v striktne vertikálnom smere. Potom vonkajší zvukovod umyté studeným príp teplá voda. Vonkajší okraj polkruhového kanálika je veľmi blízko k nemu, takže sa okamžite ochladzuje alebo zahrieva. Podľa Baraniho teórie hustota endolymfy pri zahrievaní klesá; následne jeho vyhrievaná časť stúpa a vytvára tlakový rozdiel na oboch stranách kuply; výsledná deformácia spôsobuje nystagmus (obr. 12.3; zobrazená situácia zodpovedá vyhrievaniu ľavého zvukovodu). Na základe svojej povahy sa tento typ nystagmu nazýva kalorický. Pri zahrievaní smeruje do miesta tepelného dopadu, pri ochladzovaní opačným smerom. U ľudí trpiacich vestibulárnymi poruchami sa nystagmus kvalitatívne a kvantitatívne líši od normálu. Podrobnosti o jeho testovaní sú uvedené v práci. Treba poznamenať, že kalorický nystagmus sa môže vyskytnúť v kozmickej lodi v podmienkach beztiaže, keď sú rozdiely v hustote endolymfy nevýznamné. V dôsledku toho sa na jeho spustení podieľa najmenej jeden ďalší, zatiaľ neznámy mechanizmus, napríklad priamy tepelný účinok na vestibulárny orgán.
Funkciu otolitického aparátu možno testovať pozorovaním okulomotorických reakcií pri záklone hlavy alebo pri pohybe pacienta tam a späť na špeciálnej plošine.
Vestibulárne poruchy. Silné podráždenie vestibulárneho aparátu často spôsobuje nepohodlie: závraty, vracanie, zvýšené potenie, tachykardiu atď. V takýchto prípadoch sa hovorí o kinetóza(choroba, "morská choroba"), S najväčšou pravdepodobnosťou je to dôsledok vystavenia komplexu podnetov neobvyklých pre telo (napríklad na mori): Coriolisovo zrýchlenie alebo nezrovnalosti medzi vizuálnymi a vestibulárnymi signálmi. U novorodencov a pacientov so vzdialenými labyrintmi sa kinetóza nepozoruje.
Aby sme pochopili dôvody ich výskytu, je potrebné vziať do úvahy, že vestibulárny systém sa vyvinul z hľadiska pohybu na nohách, a nie na základe zrýchlení, ktoré sa vyskytujú v moderných lietadlách. V dôsledku toho vznikajú zmyslové ilúzie, ktoré často vedú k nehodám, napríklad keď pilot prestane vnímať rotáciu alebo sa zastaví, zle vníma jej smer a podľa toho neadekvátne reaguje.
Akútna jednostranná porucha funkcia labyrintu spôsobuje nevoľnosť, vracanie, potenie atď., ako aj závraty a niekedy aj nystagmus smerujúci na zdravú stranu. Pacienti majú tendenciu padať na stranu s poruchou funkcie. Veľmi často však klinický obraz komplikované neistotou smeru závratov, nystagmu a pádu. Pri niektorých ochoreniach, ako je Meniérov syndróm, dochádza k nadmernému tlaku endolymfy v jednom z labyrintov; v tomto prípade sú prvým výsledkom podráždenia receptorov symptómy opačného charakteru, ako sú symptómy opísané vyššie. Na rozdiel od jasných prejavov akútnych vestibulárnych porúch chronická strata funkcie jedného z labyrintov kompenzované pomerne dobre. Činnosť centrálneho vestibulárneho systému môže byť prekonfigurovaná tak, aby bola znížená odozva na abnormálnu stimuláciu, najmä ak iné zmyslové kanály, ako sú zrakové alebo hmatové, poskytujú korekčnú aferentáciu. Preto sú patologické prejavy chronických vestibulárnych porúch výraznejšie v tme.
Akútne obojstranné dysfunkcia u ľudí je zriedkavá. Pri pokusoch na zvieratách sú ich symptómy oveľa miernejšie ako pri jednostrannej poruche, keďže obojstranné prerušenie aferentácie vestibulárnych jadier neovplyvňuje "symetrie" organizmu - Stav beztiaže (pri vesmírnych letoch) neovplyvňuje polkruhové kanály, ale eliminuje vplyvom gravitácie na otolity a otolitové membrány vo všetkých makulách zaujímajú polohu určenú ich vlastnými elastickými vlastnosťami. Výsledný vzor vzrušenia sa na Zemi nikdy nenachádza, čo môže viesť k príznakom kinetózy. Keď si človek zvykne na stav beztiaže, vizuálna aferentácia sa stáva dôležitejšou a úloha otolitového aparátu sa znižuje.
Každodenné rozlišovanie medzi fyzickými a biologickými aspektmi sluchu sa odráža v terminológii. "Akustika" sa týka fyzikálnych vlastností zvuku a mechanických zariadení alebo anatomických štruktúr, ktoré ovplyvňujú. Keď už hovoríme o fyziologických procesoch sluchu a ich anatomických korelátoch, používa sa termín "sluchový".
Fyzikálne vlastnosti zvukového podnetu (akustika)
Zvuk sú vibrácie molekúl (hovoríme o vibráciách superponovaných na Brownov pohyb molekúl) elastického média (najmä vzduchu), ktoré sa v ňom šíria vo forme pozdĺžnej tlakovej vlny. Takéto oscilácie média sú generované oscilačnými telesami, ako je ladička alebo reproduktor, ktoré doň prenášajú energiu a zrýchľujú molekuly, ktoré sú im najbližšie. Z toho druhého prechádza energia na molekuly umiestnené o niečo ďalej atď. Tento proces sa šíri okolo zdroja zvuku ako vlna s rýchlosťou (vo vzduchu) asi 335 m/s. V dôsledku vibrácií molekúl v médiu vznikajú zóny s väčšou alebo menšou hustotou ich balenia, kde je tlak vyšší alebo nižší ako priemer. Amplitúda jeho zmeny je tzv akustický tlak. Dá sa merať pomocou špeciálnych mikrofónov registráciou efektívnej hodnoty (pozri učebnicu fyziky) a frekvenčných vlastností, ktoré slúžia ako charakteristika zvuku. Ako každý iný, aj akustický tlak sa vyjadruje v N / m 2 (Pa), v akustike sa však zvyčajne používa porovnávacia hodnota - tzv. hladina akustického tlaku(SPL), merané v decibeloch (dB). Aby sme to dosiahli, akustický tlak p x, ktorý nás zaujíma, sa vydelí ľubovoľne zvolenou referenčnou hodnotou p 0 rovnajúcou sa 2–10 -5 N / m 2 (je blízko k hranici sluchu pre osobu) a desiatkový logaritmus kvocient sa vynásobí 20.
SPL = 20lgp x / po[db]
Logaritmická stupnica bola zvolená, pretože uľahčuje opísanie širokého rozsahu akustického tlaku v rámci počúvania. Faktor 20 sa dá ľahko vysvetliť: desiatkový logaritmus pomeru sily zvukov (I), pôvodne nazývaný „bel“ (na počesť Alexandra Bella), sa rovná 10 dB. Akustický tlak p sa však meria ľahšie ako intenzita zvuku. Pretože táto je úmerná druhej mocnine amplitúdy tlaku (I ~ p 2) a Igp 2 = 2 lgp , tento koeficient sa zavedie do rovnice. Takéto merania sa vykonávajú najmä v komunikačnej technike. Hladina akustického tlaku pre tón s akustickým tlakom 2 10 -1 N/m 2 sa napríklad vypočíta takto:
p x / po= 2• 10 –1 /2• 10 –5 =10 4, SPL= 20 1g 10 4 =20 4=80.
Takže akustický tlak 2–10 –1 N/m2 zodpovedá SPL 80 dB. Je ľahké vidieť, že zdvojnásobenie akustického tlaku zvyšuje SPL o 6 dB a zvýšenie o 10 sa rovná 20 dB. Súradnice na obr. 12.8 vľavo ilustruje vzťah medzi týmito parametrami.
V akustike sa zvyčajne špecifikuje "dB SPL", pretože dB-škála sa široko používa na opis iných javov (ako je napätie) alebo s inými konvenčnými štandardnými hodnotami. Dodatok "SPL" zdôrazňuje, že číslo sa získa z vyššie uvedenej rovnice s p o \u003d 2 10 -5 N/m2.
Pevnosťzvuk je množstvo energie, ktorá prejde jednotkovým povrchom za jednotku času; vyjadruje sa vo W/m 2 . Hodnota 10 -12 W/m 2 v rovine zvukovej vlny zodpovedá tlaku 2 10 -5 N/m 2 .
Frekvencia zvuku je vyjadrená v hertzoch (Hz); jeden hertz sa rovná jednému cyklu oscilácií za sekundu. Frekvencia zvuku je rovnaká ako frekvencia jeho zdroja, ak je tento zdroj nehybný.
Zvuk produkovaný vibráciami rovnakej frekvencie sa nazýva tón. Na obr. 12.5, A ukazuje časovú charakteristiku akustického tlaku pre tento prípad. Čisté tóny sa však v každodennom živote prakticky nikdy nenachádzajú; väčšina zvukov je tvorená superpozíciou niekoľkých frekvencií (obr. 12.5, B). Zvyčajne ide o kombináciu základnej frekvencie a niekoľkých harmonických, ktoré sú jej násobkami. Toto sú hudobné zvuky. Základná frekvencia sa odráža

Ryža. 12.5.Zmena akustického tlaku (p) v priebehu času: ALE-čistý tón; B- hudobný zvuk; IN- hluk. T- obdobie hlavnej hudobnej frekvencie; hluk nemá periódu
v období komplexnej vlny akustického tlaku (T na obr. 12.5, B). Keďže rôzne zdroje tvoria rôzne harmonické, zvuky na rovnakej základnej frekvencii sa môžu líšiť, čím sa pri hraní orchestra dosahujú bohaté odtiene zvuku. Zvuk zložený z mnohých nesúvisiacich frekvencií sa nazýva tzv hluk(obr. 12.5, IN), najmä „biely šum“, ak sú v ňom rovnako zastúpené takmer všetky frekvencie v počuteľnom rozsahu. Registráciou akustického tlaku hluku nie je možné zistiť periodicitu.
Anatomické základy sluchu; periférna časť ucha
Zvukové vlny sú posielané do sluchového systému cez externé ucho - vonkajší zvukovod - do ušný bubienok(obr. 12.6). Táto tenká, perleťová membrána oddeľuje zvukovod od stredného ucha, ktorý obsahuje aj vzduch. V dutine stredného ucha je reťaz troch pohyblivo kĺbových sluchové kostičky: malleus ( malleus ), nákovy ( incus ) A strmeň ( kroky ). "Rukoväť" paličky je pevne spojená s tympanickou membránou a základňa strmeňa (ktorá v skutočnosti vyzerá ako strmeň) zapadá do petrosálneho otvoru. oválne okno. Tu strmeň hraničí vnútorné ucho. Zvuková energia sa do nej prenáša z ušného bubienka cez kladivo, nákovu a strmeň, ktoré s ním synchrónne kmitajú. Stredoušná dutina je spojená s hltanom Eustachovou trubicou. o

Ryža. 12.6.Schéma vonkajšieho, stredného a vnútorného ucha. M - kladivo, N - nákova, C - strmeň. Šípky označujú zodpovedajúce smery pohybu tympanickej membrány (keď je zakrivená dovnútra), kĺbové spojenia medzi nákovou a strmeňom a kochleárnou tekutinou.
pri prehĺtaní sa tento priechod otvára, ventiluje stredné ucho a vyrovnáva tlak v ňom s atmosférickým tlakom. Počas zápalového procesu tu sliznice napučiavajú a uzatvárajú lumen trubice. Ak sa zmení vonkajší tlak (napríklad v lietadle) alebo sa vzduch „vypumpuje“ zo stredoušnej dutiny, vznikne tlakový rozdiel – „zapcháva uši“. Pri potápaní je tiež dôležité zvážiť tlak v tomto vzdušnom priestore; potápač sa musí pokúsiť vyrovnať vzduch v ústnej dutine („vyfúknutím uší“) alebo prehĺtaním vyrovnať sa so zvyšujúcim sa vonkajším tlakom. Ak sa to nepodarí, hrozí prasknutie bubienka.
Vnútorné ucho je umiestnené v skalnej časti spánkovej kosti spolu s orgánom rovnováhy. Kvôli svojmu tvaru sluchový orgán pomenovaný slimák ( slimák ). Pozostáva z troch paralelných kanálov zvinutých dohromady - tympanického ( scala tympani), stredná (scala media ) A vestibulárny ( scala vestibuli )schody.vestibulárny A bubnové schody prepojené cez helicotrema(obr. 12.6). Sú naplnené perilymfa, zloženie je podobné extracelulárnej tekutine a obsahuje veľa sodíkových iónov (asi 140 mmol / l). Toto je pravdepodobne ultrafiltrát plazmy. Priestory vyplnené perilymfou a cerebrospinálnou tekutinou sú vzájomne prepojené, ale ich funkčný vzťah nie je známy. V každom prípade sú mozgovomiechový mok a perilymfa veľmi podobné chemickým zložením.
stredné schody naplnené endolymfa. Táto kvapalina je bohatá na draselné ióny (približne 155 mmol/l), t.j. pripomína intracelulárne. Peri- a endolymfatické priestory slimáka sú spojené s príslušnými oblasťami vestibulárneho aparátu (obr. 12.6). Základ strmeňa v oválnom okienku prilieha k perilymfe vestibulárnej scaly; otvor sa zatvorí odkaz na zvonenie, aby tekutina nemohla preniknúť do stredného ucha. Komunikuje so základňou scala tympani ďalším otvorom. - okrúhle okno tiež uzavretá tenkou membránou, ktorá drží perilymfu vo vnútri.
Na obr. 12.7 znázorňuje priečny rez slimákom. Vestibulárna scala je oddelená od stredu Reisnerova membrána, a stred od tympanickej - hlavny (bazilárnej) membrány. Zahusťovanie, ktoré prebieha pozdĺž posledného, je Corti orgán- obsahuje receptory obklopené podpornými bunkami. Receptory sú vláskové bunky, ktoré však nesú iba stereocíliu; ich kinocília sú znížené. Rozlišovať vnútorné a vonkajšie vlasové bunky, usporiadané v jednom a troch radoch. Ľudia majú približne 3 500 vnútorných a 12 000 vonkajších vláskových buniek.
Rovnako ako vo vestibulárnom aparáte sú sekundárne senzorické bunky. Aferentné vlákna, ktoré ich inervujú, odchádzajú z bipolárnych buniek umiestnených v strede kochley špirálový ganglion; ich ostatné procesy sú posielané do centrálneho nervového systému. Asi 90 % nervových vlákien špirálového ganglia končí vo vnútorných vláskových bunkách, z ktorých každá tvorí kontakty s mnohými z nich; zvyšných 10 % inervuje oveľa početnejšie vonkajšie vláskové bunky. Na dosiahnutie všetkých sa tieto vlákna silne rozvetvujú, hoci receptory inervované jedným vláknom sú umiestnené blízko seba. Celkovo je v sluchovom nerve približne 30 000 – 40 000 aferentných vlákien. Eferenty sú vhodné aj pre Cortiho orgán, ktorého funkčný význam je nejasný, hoci je známe, že môžu inhibovať aktivitu aferentácie.
Nad Cortiho orgánom leží tectorial (krycia vrstva) membrána - rôsolovitá hmota spojená so sebou a s vnútornou stenou slimáka. Táto membrána oddeľuje úzky priestor naplnený tekutinou pod ňou od endolymfy scala media nad ňou. Konce stereocilia vonkajších vláskových buniek sú spojené so spodným povrchom tektoriálnej membrány. Pravdepodobne sú s ňou v kontakte aj riasinky vnútorných vláskových buniek, aj keď oveľa menej tuho; táto otázka ešte nie je definitívne objasnená.
Na vonkajšej strane sa nachádza stredné schodisko cievny pásik ( stria vascularis ) je oblasť s vysokou metabolickou aktivitou a dobrým prekrvením, čomu zodpovedá aj jej názov. Hrá dôležitú úlohu v dodáva energiu slimákom a regulácia zloženia endolymfy. Rôzne iónové pumpy, vrátane draslíka, udržujú stálosť iónového média a jeho pozitívny potenciál. Je známe, že niektoré diuretiká (látky zvyšujúce močenie) majú ototoxické účinky. vedľajší účinok a môžu viesť k hluchote, pretože ovplyvňujú iónové pumpy cievnych strií. Tie isté látky blokujú iónové pumpy v epiteli renálnych tubulov (pozri časť 30.4), ktoré sú zodpovedné za reabsorpciu solí. Je zrejmé, že niektoré mechanizmy transportu iónov sú v oboch prípadoch podobné.
Hranice sluchu . Aby bol zvuk počuteľný, musí byť prekročená určitá hladina akustického tlaku (SPL). Tento prah (obrázok 12.8) je závislý od frekvencie; ľudské ucho najcitlivejšie v rozsahu 2000–5000 Hz. Okrem toho sú na dosiahnutie prahu potrebné výrazne vyššie SPL.

Ryža. 12.7. Schéma rezu vnútorného ucha. Hore je poloha slimáka, špirálového ganglia a sluchového nervu. Nižšie sú uvedené najdôležitejšie prvky jednej z cievok slimáka a jej lymfatických priestorov. Zloženie subtektoriálnej lymfy nebolo presne stanovené. Ukázané sú aj priestorové spojenia medzi tektoriálnou membránou a receptorovými bunkami Cortiho orgánu.
Objem . Tón ktorejkoľvek frekvencie nám pri prekročení prahu počutia znie so zvyšujúcim sa akustickým tlakom hlasnejšie. Vzťah medzi fyzickou hodnotou ultrazvuku a subjektívne vnímaným objem možno kvantifikovať. Inými slovami, človek môže zistiť nielen to, či daný tón počuje, ale aj to, či vníma dva po sebe nasledujúce tóny rovnakej alebo rozdielnej frekvencie ako rovnako nahlas alebo v tomto smere odlišná. Napríklad jeden po druhom sú prezentované testovacie a referenčné tóny s frekvenciou 1 kHz a subjekt je vyzvaný, aby potenciometrom upravil hlasitosť druhého zvuku tak, aby ho vnímal ako
rovnaký ako predchádzajúci. Hlasitosť akéhokoľvek zvuku je vyjadrená v fónoch - SPL tónu s frekvenciou 1 kHz s rovnakou hlasitosťou. Ak sa teda vo vyššie uvedenom príklade subjektívny vnem vyrovná na 70 dB, potom je hlasitosť testovaného tónu 70 fónov. Pretože sa ako štandard používa 1 kHz, sú tu hodnoty decibelov a phonov rovnaký(obr. 12.8). Na obr. 12.8 tiež ukazuje krivky rovnakej počuteľnosti, postavené na priemernej odpovedi mladých zdravých jedincov (veľká medzinárodná vzorka). Všetky tóny na každej krivke sa považujú za rovnako hlasné bez ohľadu na ich frekvenciu. Takéto krivky sa nazývajú izofóny. Tu uvedený prah

Ryža. 12.8.Rovnaké krivky hlasitosti (izofóny) podľa nemeckej normy DIN 45630. Na osi y sú vľavo vynesené ekvivalentné hodnoty akustického tlaku a SPL. červená je označená oblasť reči (pozri text)
krivka je tiež izofóna, pretože všetky jej tóny sú vnímané ako rovnako hlasné, t.j. sotva počuteľné. Priemerný prah sluchu u zdravého človeka je 4 von, aj keď, samozrejme, sú možné odchýlky od tejto hodnoty v oboch smeroch.
Prah rozlišovania intenzity zvuku . Keďže škála pozadia je založená na subjektívnom vnímaní, je zaujímavé zistiť, nakoľko je presná, t.j. ako rozdielne sa musia líšiť akustické tlaky dvoch tónov (ktoré pre zjednodušenie môžu mať rovnakú frekvenciu), aby bola ich hlasitosť vnímaná nerovnomerne. V experimentoch na meranie prah intenzity zvuku tento rozdiel bol veľmi malý. Na prahu počutia sú dva tóny rovnakej frekvencie vnímané ako nerovnako hlasné, keď sa ich SPL líšia o 3–5 dB. Pri sile zvuku asi 40 dB nad prahom počutia klesá táto hodnota na 1 dB.
Samotná škála pozadia nehovorí nič o subjektívnom zvýšiť hlasitosť so zvyšujúcim sa ultrazvukom. Vychádza len zo slov subjektu, ktorý určuje, kedy sa mu hlasitosť skúšobných a referenčných tónov zdá rovnaká; koľko objem sa mu zmenil, v tomto prípade sa to vôbec neskúma. Zároveň je zaujímavý vzťah medzi ním a akustickým tlakom, keďže pri posudzovaní zdraviu škodlivého hluku treba brať do úvahy zmeny vnímanej hlasitosti. Na určenie tohto vzťahu bol subjekt požiadaný, aby upravil testovací tón pri frekvencii 1 kHz tak, aby vyzeral v n krát hlasnejšie (napríklad 2 alebo 4 krát) ako referencia s rovnakou frekvenciou a 40 dB SPL. Na základe takto získaných ultrazvukov je možné kvantifikovať intenzitu vnemu; táto jednotka objemu sa nazýva sójové bôby. Hlasitosť tónu, ktorý znie 4-krát hlasnejšie ako štandardný pre človeka, je 4 son, o polovicu tichšia ako 0,5 sona atď.
Ukázalo sa, že pri SPL nad 30 dB je pocit hlasitosti spojený s akustickým tlakom s mocninovým zákonom s exponentom 0,6 pri frekvencii 1 kHz (Stevensova silová funkcia; pozri).
Inými slovami, pri frekvencii 1 kHz a SPL nad 30 dB sa pocit hlasitosti zdvojnásobí, keď sa SPL zvýši o 10 dB. Keďže zdvojnásobenie akustického tlaku zodpovedá zvýšeniu SPL o 6 dB, vnímaná hlasitosť sa nezdvojnásobuje paralelne – akustický tlak by sa musel takmer strojnásobiť. Preto, keďže I ~ p 2, aby sa zdvojnásobila subjektívna hlasitosť, sila zvuku sa musí zvýšiť 10-krát. To znamená, že hlasitosť desiatich hudobných nástrojov hrajúcich v rovnakom tóne s rovnakým SPL je len dvakrát vyššia ako hlasitosť jedného z nich.
Pretože pre každý je hlasitosť v fónoch podľa definície odvodená od zvuku tónu 1 kHz, hlasitosť akéhokoľvek tónu v synoch sa môže vypočítať z počtu fónov v ňom a krivky hlasitosti tónu 1 kHz. Na technické merania škodlivého hluku sa používa zjednodušený postup, ktorý udáva približné hodnoty hlasitosti v fónoch.
Prístroje na meranie SPL a úrovne hlasitosti . Ako spomenuté vyššie, izofóny boli získané v psychofyzikálnych experimentoch. Preto nie je možné určiť hlasitosť v pozadí fyzikálnymi metódami, ako sa to robí pri meraní pomocou vhodných mikrofónov a zosilňovačov akustického tlaku. Aby ste aspoň približne zmerali úroveň hlasitosti, môžete použiť rovnaké prístroje s frekvenčnými filtrami, ktoré svojimi charakteristikami približne zodpovedajú prahu počutia alebo iné izofóny, teda prístroje s takmer rovnakou nerovnakou citlivosťou na rôzne frekvencie ako ľudské ucho: menej citlivé na nízke a vysoké frekvencie. Existujú tri takéto medzinárodné hodnotenia filtrov – A, B a C. Pri citovaní výsledkov merania uveďte, ktorý z nich je použitý, pridaním príslušného písmena k hodnote decibelov, napríklad 30 dB (A), čo znamená približne 30 fónov. Odozva filtra A sleduje prahovú krivku a teoreticky by sa mala použiť len pri nízkych hladinách zvuku, ale pre jednoduchosť sa teraz takmer všetky výsledky uvádzajú vo videodB(A), aj keď to predstavuje ďalšiu chybu. Rovnaká stupnica sa používa pri meraní škodlivých zvukov, aj keď prísne vzaté, v tomto prípade by sa mala použiť stupnica spánku. Napríklad hlučnosť auta na voľnobeh je asi 75 dB(A).
zvuková trauma . Ak prudko zvýšite ultrazvuk, nakoniec dôjde k pocitu bolesť v ušiach. Experimenty ukázali, že to vyžaduje úroveň hlasitosti približne 130 fónov. Navyše zvuk takejto intenzity spôsobuje nielen bolesť, ale aj reverzibilnú stratu sluchu (dočasné zvýšenie prahu sluchu) alebo pri dlhšom ožiarení jeho nevratnú stratu (pretrvávajúce zvýšenie prahu sluchu, zvuková trauma). V tomto prípade sú poškodené zmyslové bunky a narušená mikrocirkulácia v slimáku. Poškodenie zvuku môže nastať aj pri dostatočne dlhom vystavení oveľa slabším zvukom s intenzitou aspoň 90 dB (A).
Osoby pravidelne vystavené takýmto zvukom sú vystavené riziku straty sluchu; a mali by používať bezpečnostné zariadenia (slúchadlá, štuple do uší). Ak neprijmete opatrenia, strata sluchu sa vyvinie v priebehu niekoľkých rokov.
Subjektívne reakcie na hluk . Okrem zvukovej traumy, t. j. objektívne pozorovaného poškodenia vnútorného ucha, môže zvuk spôsobiť aj niektoré nepríjemné pocity subjektívneho charakteru (niekedy sprevádzané objektívnymi príznakmi – zvýšená krvný tlak, nespavosť atď.). Nepohodlie spôsobené hlukom do značnej miery závisí od psychologického postoja subjektu k zdroju zvuku. Napríklad obyvateľa domu môže veľmi otravovať hra na klavíri o dve poschodia vyššie, hoci úroveň hlasitosti je objektívne nízka a ostatní obyvatelia nemajú žiadne sťažnosti. Je ťažké nájsť všeobecné pravidlá, ako predísť pre človeka nepríjemným zvukom, a právne predpisy platné v tomto smere sú často len nevyhovujúcimi kompromismi.
Hranice oblasti sluchu a reči . Počuteľnosť tónu, ako je znázornené na obr. 12.8 závisí od jeho frekvencie a akustického tlaku. Mladý zdravý človek rozlišuje frekvencie od 20 do 16000 Hz (16 kHz). Frekvencie nad 16 kHz sú tzv ultrazvuk, a pod 20 Hz - infrazvuk. Sluchové limity pre ľudí sú teda 20 Hz–16 kHz a 4–130 fónov. Na obr. 12.8 sluchová zóna umiestnené medzi hornou a dolnou krivkou. Frekvencie a sily zvuku charakteristické pre reč sú v strede tejto oblasti (na obrázku sú vytieňované červenou farbou); korešpondujú rečová zóna. Na zabezpečenie primeraného porozumenia reči musia komunikačné systémy (napr. telefón) prenášať frekvencie v rozsahu najmenej 300 Hz až 3,5 kHz. Citlivosť na vysoké frekvencie postupne s vekom klesá (tzv starecká hluchota).
Prah frekvenčnej diskriminácie . Z každodennej skúsenosti je známe, že tóny sa líšia nielen hlasitosťou, ale aj výška,čo koreluje s ich frekvenciou. Hovorí sa, že tón je vysoký, ak je jeho frekvencia vysoká a naopak. Schopnosť človeka rozlíšiť výšky postupne počutých tónov je prekvapivo vysoká. V optimálnej oblasti okolo 1 kHz prah rozlišovania frekvencie je 0,3 %, t.j. približne 3 Hz.
Hudobným zvukom s viacerými frekvenciami možno tiež priradiť špecifickú výšku tónu; zvyčajne sa považuje za rovnaký ako čistý tón so základnou zvukovou frekvenciou. Obvyklá hudobná stupnica je rozdelená na oktávy;
zvuky rovnakého mena v susedných oktávach sa líšia frekvenciou o polovicu. Temperovaná oktáva je rozdelená na 12 krokov, z ktorých každý sa od nasledujúceho líši vo frekvencii 1,0595-krát. Tento rozdiel je v podstate vyššie uvedený prah rozlišovania frekvencie. Na rozlíšenie dvoch súčasne znejúcich čistých tónov je však ich frekvenčný rozdiel oveľa väčší, ako keď nasledujú jeden za druhým. Je zrejmé, že na to musia byť dve súčasne stimulované oblasti vnútorného ucha oddelené určitou minimálnou vzdialenosťou.
Odtiaľ pochádza pojem „kritické frekvenčné pásmo“. Napríklad sa zistilo, že sluchový systém nie je schopný rozlíšiť čisté tóny v rámci tretiny oktávy (toto je kritické pásmo); spájajú sa a vytvárajú pocit jedného zvuku. S nárastom počtu komponentov, ktoré tvoria zvuk v tomto frekvenčnom rozsahu, sa zvyšuje iba subjektívna hlasitosť, ale výška vnímaná osobou sa nemení. Zvuková energia v kritickom pásme je teda sčítaná, čo spôsobuje jediný pocit.
Kritické pásmo je prekvapivo široké: nie je možné rozlíšiť medzi dvoma znejúcimi čistý tóny oddelené takmer tretinou oktávy. V prípade zmiešaných tónov je situácia samozrejme iná: je ľahké určiť, kedy sú súčasne stlačené dve susedné klávesy klavíra, pretože nie všetky harmonické, ktoré prekrývajú základnú frekvenciu každej noty, sú zahrnuté v jednom kritickom pásme.
Približne 24 kritických pásiem sa zmestí do hraníc ľudského sluchu. Táto problematika sa podrobnejšie zaoberá v prácach.
Keď zaznejú dva tóny súčasne, zmenia sa prahy sluchu oboch. Napríklad na pozadí konštantného tónu s frekvenciou 500 Hz s SPL 80 dB, ostatné tóny s intenzitou zvuku zodpovedajúcou prahu ich počuteľnosti na obr. 12.8 nie sú akceptované. Pre ich počuteľnosť je potrebný oveľa vyšší SPL, konkrétne asi 40 dB pre frekvenciu 1 kHz. Tento jav sa nazýva maskovanie. Má to veľký praktický význam, pretože v každodennom živote môžu byť dôležité akustické informácie, ako napríklad rozhovor, tak maskované hlukom v pozadí, že sa stávajú úplne nezrozumiteľné. Psychoakustické javy sú podrobnejšie opísané v prácach.
Ako už bolo spomenuté, ušný bubienok vibruje zvukom a odovzdáva svoju energiu vo vzduchu pozdĺž kostnej perilymfy vestibulárnej scaly.
Zvuk sa potom šíri do tekutého média vnútorného ucha; kým väčšina jeho energie odrážal od rozhrania medzi médiami, keďže sa líšia akustickým odporom (impedanciou). ale osteotympanického aparátu stredné ucho navzájom „ladí“ impedancie oboch médií, čím výrazne znižuje stratu spätného toku. V prvej aproximácii to možno porovnať s pôsobením šošovky fotoaparátu, ktorá znižuje odraz svetla na rozhraní vzduch-sklo. Impedančné prispôsobenie zaisťujú dva mechanizmy. Po prvé, plocha tympanickej membrány je oveľa väčšia ako na spodnej časti strmeňa a keďže tlak je priamo úmerný sile a nepriamo k ploche, je vyšší v oválnom okienku ako na bubienku. Po druhé, dochádza k dodatočnému zvýšeniu tlaku v dôsledku zmeny ramien páky vytvorených reťazou kostí. Celý systém teda funguje ako zosilňovací elektrický transformátor, hoci v procese pôsobia aj iné faktory – hmotnosť a elasticita prepojených kostí, ako aj zakrivenie a vibračné vlastnosti bubienka. Mechanizmus prispôsobenia impedancie zlepšuje sluch o 10-20 dB; v závislosti od frekvencie je to ekvivalentné 2- až 4-násobnému zvýšeniu vnímanej hlasitosti. Vodivé vlastnosti tympano-kostného aparátu sú určené frekvenciou. Najlepší prenos je pozorovaný v strede frekvenčného rozsahu počuteľnosti, čo čiastočne určuje tvar krivky, ktorá charakterizuje jej prah.
Pocit zvuku sa vyskytuje aj vtedy, keď sa oscilujúci predmet, ako napríklad ladička, položí priamo na lebku; v tomto prípade je hlavná časť energie prenášaná kosťami týchto kostí (tzv kostné vedenie). Ako bude ukázané v ďalšej časti, excitácia receptorov vo vnútornom uchu vyžaduje pohyb tekutiny, ako je pohyb spôsobený vibráciami strmeňa, keď sa zvuk šíri vzduchom. Zvuk prenášaný kosťami spôsobuje tento pohyb dvoma spôsobmi. Po prvé, vlny kompresie a riedenia, ktoré prechádzajú cez lebku, vytlačia tekutinu z objemného vestibulárneho labyrintu do slimáka a potom späť (teória kompresie). Po druhé, hmotnosť tympano-kostného aparátu a s tým spojená zotrvačnosť vedie k oneskoreniu jeho oscilácií od oscilácií charakteristických pre kosti lebky. V dôsledku toho sa strmeň pohybuje vzhľadom na skalnú kosť, čím vzrušuje vnútorné ucho (teória hmotnosti a zotrvačnosti).
V bežnom živote nie je kostné vedenie také výrazné. Pokiaľ sa vám vlastný hlas nahraný na magnetofone (najmä v nízkofrekvenčnom pásme) nezdá na nerozoznanie, keďže v živej reči sa časť energie prenáša do ucha cez kosti. Kostné vedenie je však široko používané v diagnostike.
Svaly stredného ucha (m. tensor tympani, m. stapedius ) sú pripevnené ku kladivu a strmeňovi, resp. Keď sú vystavené zvuku, ich reflexná kontrakcia tlmí prenos, keď sa mení impedancia stredného ucha. Tento mechanizmus nechráni pred zvukmi nadmernej hlasitosti, aj keď sa o tejto možnosti diskutovalo. Funkčný význam reflexov stredného ucha zostáva nejasný.
Procesy sluchu vo vnútornom uchu
mechanické javy. Keď zvuk spôsobí rozkmitanie strmeňa, odovzdá svoju energiu perilymfe vestibulárnej šupiny (obr. 12.6). Keďže tekutina vo vnútornom uchu je nestlačiteľná, musí tam byť nejaká štruktúra
vyrovnávanie tlaku. Toto je okrúhle okno. Jeho membrána sa ohýba v smere opačnom k pohybu strmeňa. Tá zároveň vytiahne z pokoja bazálnu časť strednej skaly, ktorá je najbližšie k nej, spolu s Reisnerovými a bazilárnymi membránami, ktoré ju pokrývajú, a osciluje hore a dole v smere vestibulárnej, potom k bubienkovej šupke — Pre jednoduchosť budeme odteraz volať stredné schodisko s jeho membránami endolymfatický kanál. Posun jeho základne generuje vlnu, ktorá sa šíri od strmeňa k helikotrému ako pozdĺž napnutého lana. Na obr. 12,9, A sú znázornené dva stavy takejto vlny (endolymfatický kanál je znázornený jednou čiarou). Keďže zvuk neustále kmitá strmeňom, tzv putujúce vlny(cm). Tuhosť bazilárnej membrány od strmeňa po helikotrému klesá, takže rýchlosť šírenia vĺn sa postupne znižuje a ich dĺžka sa zmenšuje. Z rovnakého dôvodu sa ich amplitúda najskôr zväčší (obr. 12.9), pričom je oveľa väčšia ako v blízkosti strmeňa, ale pod vplyvom tlmiacich vlastností tekutinou naplnených kanálikov vnútorného ucha sa čoskoro potom zníži na nulu, zvyčajne pred helikotrémou. Niekde medzi bodmi výskytu vlny a jej útlmu sa nachádza úsek, kde je jej amplitúda maximálna (obr. 12.9). Toto maximum amplitúdy závisí od frekvencie: čím je vyššia, tým je bližšie k strmeňu, čím nižšia, tým ďalej. Výsledkom je, že maximum amplitúdy každej frekvencie v rozsahu počuteľnosti zodpovedá špecifickému úseku endolymfatického kanála (bazilárnej membrány). Volá sa frekvenčný rozptyl. Senzorické bunky sú najviac excitované tam, kde je amplitúda oscilácie maximálna, takže na rôzne bunky pôsobia rôzne frekvencie. (teória miesta).
Vyššie opísané pohyby vĺn a najmä polohu maxima amplitúdy možno pozorovať pomocou Mössbauerovej metódy, kapacitného senzora alebo interferometrických metód. Je pozoruhodné, že aj maximálna amplitúda vĺn je extrémne malá. Pre zvuk na prahu počutia je priehyb membrány len asi 10–10 m (približne priemer atómu vodíka!). Ďalším dôležitým bodom je prísna lokalizácia amplitúdového maxima: rôzne časti bazilárnej membrány sú veľmi jasne „naladené“ na určitú frekvenciu, ak je slimák úplne neporušený. Ak sa poškodí (napríklad pri miernej hypoxii), amplitúda oscilácií sa zníži a takéto jemné doladenie sa stratí. Inými slovami, bazilárna membrána nielen pasívne osciluje; aktívne procesy zabezpečuje frekvenčne špecifický zosilňovací mechanizmus.
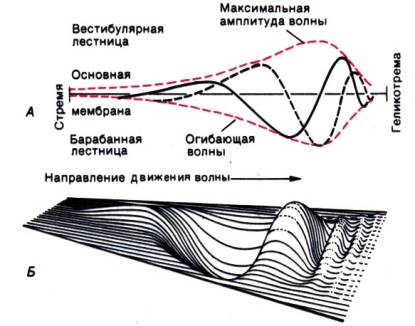
Ryža. 12.9. ALE. Schéma postupujúcej vlny v dvoch časových bodoch. Obálka ukazuje svoju maximálnu amplitúdu pri konštantnej frekvencii v rôznych častiach kochley. B. 3D rekonštrukcia vĺn
Transformačné procesy vo vlasových bunkách . Ako už bolo spomenuté v predchádzajúcej časti, v dôsledku mechanických vlastností slimáka určitá zvuková frekvencia spôsobuje kmity bazilárnej membrány s amplitúdou dostatočnou na vybudenie zmyslových buniek len na jednom, prísne obmedzenom mieste. Od bazilárnych a tektoriálnych membrán pohybujúce sa voči sebe navzájom na mihalnice pôsobí šmyková sila, jednak pri ich priamom kontakte s tektoriálnou membránou, jednak v dôsledku pohybu subtektorálnej lymfy; v oboch prípadoch ich ohnutie slúži ako adekvátny stimul pre sluchové receptory (ako pri vestibulárnych receptoroch).
Toto ohýbanie začína proces konverzie(transdukcia): mikroskopická mechanická deformácia mihalníc vedie k otvoreniu iónových kanálov v membráne vláskových buniek a následne k ich depolarizácii. Jeho predpokladom je prítomnosť endokochleárny potenciál. Mikroelektródové merania ukázali, že endolymfatický priestor má kladný (približne +80 mV) náboj v porovnaní so scala vestibularis a inými extracelulárnymi priestormi tela. Cievny pruh a Cortiho orgán nesú záporný náboj (~ -70 mV; Obr. 12.10). Potenciály zaznamenané v Cortiho orgáne pravdepodobne zodpovedajú intracelulárnym potenciálom vlasových a podporných buniek. Pozitívny endokochleárny potenciál zabezpečujú energeticky závislé procesy v cievnom pruhu. Posun mihalníc počas stimulácie mení odpor membrány vláskových buniek v dôsledku otvorenia iónových kanálov. Pretože medzi

Ryža. 12.10.Konštantné potenciály kochley

Ryža. 12.11.Kochleárny mikrofonický potenciál (MP) a akčný potenciál sluchového nervu (CAP) zaznamenaný v okrúhlom okienku pri zvuku kliknutia
endolymfatického priestoru a ich vnútrobunkového prostredia je výrazný potenciálový rozdiel (najmenej 150 mV), synchrónne so stimulom vznikajú lokálne iónové prúdy, ktoré menia membránový potenciál vláskových buniek, t.j. generujú receptorový potenciál (tzv. hypotéza batérie). Registrácia je náročná, ale možná. Jednoduchšie je však zaznamenať mikroelektródy v blízkosti receptorov v scala tympani alebo na okrúhlom okienku. mikrofónny potenciál kochley(obr. 12.11).
Je podobné výstupnému napätiu mikrofónu a presne odráža zmeny akustického tlaku. Páskový záznam reči, ktorý vznikol napojením pokusného zvieraťa na mikrofónny potenciál, je celkom čitateľný. Pôvod tohto potenciálu je nejasný; počiatočný predpoklad, že pozostáva z extracelulárne zaznamenaných zložiek receptorových potenciálov vláskových buniek, už nie je celkom prijateľný. Ako ukazujú intracelulárne vodiče z vnútorných a vonkajších vláskových buniek, hoci generujú receptorové potenciály, pri vysokej frekvencii stimulov sa zaznamenáva iba konštantné napätie: membránový potenciál vláskových buniek sa nemení synchrónne s vysokofrekvenčným zvukom. Potenciál mikrofónu:
1) je synchrónny so zvukovým podnetom prakticky bez latentnej periódy;
2) je zbavená refraktérnej periódy;
3) bez merateľného prahu;
4) nepodlieha únave; tie. sa vo všetkých ohľadoch líši od nervového akčného potenciálu.
Depolarizácia vláskových buniek spôsobuje uvoľnenie mediátora (prípadne glutamátu) z ich bazálnej časti, ktorý excituje aferentné nervové vlákna. Keď sa v blízkosti ucha ozve cvaknutie (krátky tlakový impulz), vlákna sluchového nervu sa synchrónne aktivujú a z okrúhleho okienka môžete okrem mikrofónu zaznamenať aj zložený akčný potenciál. Dlhšie zvuky spôsobujú asynchrónne impulzy, ktoré sa nesčítavajú do samostatných akčných potenciálov. Na obr. 12.11 ukazuje kochleárny mikrofónny potenciál (MCP) a zložený akčný potenciál (CAP) vyvolaný kliknutím. Boli zaznamenané u mačiek, ale môžu byť zaznamenané aj u ľudí, keď sa elektróda na diagnostické účely prevlečie cez bubienok a privedie k okrúhlemu okienku.
Zvukové kódovanie vo vláknach sluchového nervu .
V kochleárnom nerve je 90 % aferentných vlákien myelinizovaných a pochádzajú z vnútorných vláskových buniek. Každý kontaktuje len jedného z nich, t.j. s veľmi malou časťou slimáka. Tieto vlákna sú dostatočne hrubé na to, aby v nich pomocou mikroelektród registrovali akčné potenciály a študovali reakciu na zvukovú stimuláciu (vlákna vyčnievajúca z vonkajších vláskových buniek sú na to príliš tenké). Keďže každý úsek slimáka zodpovedá určitej frekvencii, každé z týchto vlákien je najviac vzrušené tým vlastným charakteristická frekvencia zvuk a iné frekvencie sa neaktivujú vôbec alebo sa aktivujú až so zvýšením akustického tlaku. To sa odráža na obr. 12.12, ktorý ukazuje graf prahu vnímania versus frekvencia stimulov pre dve rôzne vlákna. Kritériom pre stanovenie prahu je určitý nárast aktivity nad jej spontánnu úroveň. Každé vlákno je vypálené pri hodnotách frekvencie a intenzity v rámci oblasti vytieňovanej na obrázku. Na krivke, ktorá ho ohraničuje prahová frekvenčná odozva nápadná je úzka, špicatá nízkoprahová oblasť a široká vysokoprahová oblasť. Charakteristiky frekvenčného prahu odrážajú

Ryža. 12.12.Schematické frekvenčno-prahové krivky dvoch aferentných vlákien sluchového nervu (a, b) s rôznymi charakteristickými frekvenciami (HF). Krivka v je typická pre vlákno s patologickými zmenami spôsobenými poškodením vnútorného ucha.
rozloženie frekvenčných maxím na hlavnej membráne. Odozva jedného vlákna na stimul, vyjadrená ako taká krivka, je spektrálna analýza zvuku. Ak je v ňom niekoľko rôznych frekvencií, aktivuje sa niekoľko skupín nervových vlákien. Trvanie zvukový podnet je zakódovaný trvaním nervovej činnosti, a intenzita-jejúrovni. S nárastom akustického tlaku sa zvyšuje aj frekvencia neurónových impulzov (až do určitej hranice, po ktorej nastáva saturácia). Pri veľmi vysokom tlaku sa navyše aktivujú aj susedné vlákna, ktoré boli predtým v pokoji. Takýto proces je znázornený na obr. 12,12; obe vlákna sú excitované, ak zvuk zodpovedá oblasti prekrytia ich frekvenčno-prahových charakteristík. Takže na úrovni primárnej aferentácie sa zvukový podnet rozkladá na frekvenčné zložky. Každý z nich vzrušuje príslušné nervové vlákna. Na vyšších úrovniach sluchového traktu sa neuróny môžu správať inak.
Pri poškodení slimáka klesá citlivosť a frekvenčná selektivita aferentných vlákien (obr. 12.12). Receptorový potenciál vnútorných vláskových buniek sa tiež mení a ako je uvedené vyššie, to isté sa deje s mechanickými vibračnými vlastnosťami hlavnej membrány. Možno predpokladať, že tieto bunky určujú správanie týchto buniek a vlákien, ale samotné závisia od procesu aktívneho mechanického spevnenia, za ktorý sú pravdepodobne zodpovedné vonkajšie vlasové bunky. Podľa existujúcej hypotézy sú najskôr stimulované zvukom a vytvárajú dodatočnú vibračnú energiu rovnakej frekvencie. Potom sa prenesie do vnútorných vlasových buniek. Ak je to pravda, hovoríme o akomsi hybride medzi zmyslovou bunkou a generátorom mechanickej energie.
Mnoho otázok tu ešte nebolo zodpovedaných, ale schopnosť kochley produkovať a analyzovať zvukovú energiu je nepopierateľná. Zvuk generovaný v slimáku je možné merať aj mimo ušného bubienka. Tieto procesy sú veľmi často porušované rôznymi zraneniami.
Vonkajšie vláskové bunky sú vhodné ako aktívne zosilňovače, pretože obsahujú kontraktilné proteíny a slúži im vysoko vyvinutá eferentná sieť. Navyše ich aferentácia zjavne nie je podstatná pre prenos informácií do mozgu.
Kódovanie zvukových frekvencií podľa princípu lokalizácie receptora bolo diskutované vyššie. Druhý typ kódovania informácií v sluchovom nerve je nasledujúci. Tóny do 5 kHz zvyčajne spôsobujú neurónové impulzy v sluchovom nerve iba počas určitých fáz zvukového cyklu. Výsledkom je, že časová štruktúra stimulu (napríklad perióda T na obr. 12.5, b) reprezentované skupinami akčných potenciálov prenášaných do CNS pozdĺž sluchového nervu vo vhodných časových bodoch. Mozog je zjavne schopný vyhodnotiť časovú štruktúru impulzov a určiť základnú frekvenciu zvuku (tzv analýza periodicity). Zvlášť jasné dôkazy o tom boli získané priamou elektrickou stimuláciou sluchového nervu pacientov trpiacich hluchotou; periodické podnety boli spracované tak, aby bolo cítiť tón určitej výšky, čo ukazuje skutočný význam analýzy periodicity pre sluch.
Anatómia sluchového traktu Veľmi zjednodušený diagram je znázornený na obr. 12.13. Pre jednoduchosť je znázornená len cesta z ľavého ucha. Šípky označujú synapsie s neurónmi vyššieho rádu. Aby nedošlo k preťaženiu kresby, vynechávajú sa opakujúce sa kolaterály a interkalárne neuróny, aj keď takéto spojenia sú v sluchovom systéme celkom bežné.
Primárne aferentné vlákno sa rozdvojí a pošle jeden proces do ventrálny a druhý k dorzálne kochleárne (kochleárne) jadrá. ich jemná štruktúra(najmä chrbtová) je veľmi zložitá. Ventrálny trakt (z ventrálneho jadra) vedie (čiastočne cez jadro trapézového tela) k ipsi- a kontralaterálnym olivárnym komplexom, ktorých neuróny tak prijímajú signály z oboch uší. Práve táto nervová úroveň umožňuje porovnávať akustické signály prichádzajúce z dvoch strán tela (k tomuto procesu porovnávania sa vrátime nižšie). Dorzálny trakt (od

Ryža. 12.13.Veľmi zjednodušená schéma sluchového traktu (len pre ľavé ucho). Na demonštráciu binaurálnych interakcií v hornej olive sú zobrazené aj spojenia. správny ventrálne kochleárne jadro. Odstredivé spôsoby vynechané
dorzálne jadro) prechádza na opačnú stranu tela a ide do jadro laterálneho lemnisku (laterálna slučka). Vzostupné procesy buniek olivarového komplexu sú ipsi- aj kontralaterálne. Po synaptickom prepnutí v jadre laterálnej slučky prechádza sluchová dráha colliculus inferior kvadrigemina a mediálne genikulárne telo k primárnej sluchovej kôre, pokrývajúci priečny temporálny gyrus hornej časti spánkových lalokov (Geshlov gyrus). Táto zóna zodpovedá poľu 41 podľa Brodmana; väčšina je ukrytá v hlbinách Sylvianskej brázdy. Primárna sluchová kôra susedí s ďalšími projekčnými oblasťami sluchového systému, nazývanými sekundárna sluchová kôra (pole 42 podľa Brodmanna). Prekortikálny sluchový trakt teda pozostáva z najmenej piatich alebo šiestich neurónov a keďže ďalšie synaptické prepínače a rekurentné kolaterály na obr. 12.13 nezobrazené, sú možné dlhšie reťaze. Toto je podrobnejšie popísané v prac. Nakoniec, okrem aferentných dráh, sluchový systém zahŕňa aj odstredivé eferentné vlákna, tiež neznázornené na obr. 12.13.
Excitácia centrálnych neurónov sluchového systému . Zatiaľ čo primárne aferenty sluchového nervu sú excitované čistými tónmi, teda veľmi jednoduchými zvukovými podnetmi, neuróny vyšších úrovní toho spravidla nie sú schopné. Vo ventrálnom kochleárnom jadre stále sa správajú ako primárne neuróny. Čistá

Ryža. 12.14.Aktivita štyroch neurónov dorzálneho kochleárneho jadra ako odpoveď na pôsobenie charakteristického frekvenčného tónu počas 50 ms (so zmenami). Autor: abscisa – čas; na os y počet akčných potenciálov
tóny nadprahovej intenzity vždy spôsobujú ich excitáciu; majú úzke, špicaté krivky frekvenčného prahu a krátke latentné periódy. Avšak už v dorzálne kochleárne jadro obrázok je úplne iný. Hoci aj tu väčšina neurónov vystreľuje čistými tónmi, typy ich odpovedí sa značne líšia. Ako príklad na obr. 12.14 ukazuje reakcie rôznych vlákien vychádzajúcich z tohto jadra: v každom prípade bol prezentovaný tón trvajúci 50 ms s frekvenčnou charakteristikou tejto bunky. Neurón na obr. 12.14, ALE sa správa ako primárny aferent, zatiaľ čo správanie ostatných je podstatne odlišné. V niektorých z nich môže zvuk spôsobiť brzdenie; iné sú excitované presne definovanými frekvenciami a sú brzdené miernou odchýlkou od nich. Existujú aj neuróny, ktoré reagujú zvláštnym spôsobom na zvuky premenlivej frekvencie (tzv. frekvenčne modulované tóny), hoci reagujú aj na čisté tóny. Anatomickým základom takéhoto komplexného správania sú kolaterálne spojenia, z ktorých niektoré sú excitačné a iné inhibičné.
Funkčná hodnota z toho všetkého je zrejmé, že neuróny reagujú obzvlášť jasne na určité vlastnosti zvukového podnetu, čo prispieva k rozpoznávaniu obrazu už pri najnižšia úroveň traktu. Na vyšších úrovniach sa špecifickosť ich odpovede postupne zvyšuje.
Čím ďalej od kochley pozdĺž sluchového traktu, tým zložitejšie zvukové charakteristiky sú potrebné na aktiváciu neurónov. Mnohé bunky vôbec nereagujú na čisté tóny. IN spodný colliculus kvadrigemina, napríklad existujú bunky, ktoré reagujú len na frekvenčne modulované tóny so špecifickým smerom a stupňom modulácie. Ostatné neuróny tu reagujú len na amplitúdovo modulované (tj premenlivú intenzitu) tóny. A v tomto prípade musí mať modulácia často určité vlastnosti, inak nespôsobí budenie.
Vo všeobecnosti môžeme povedať, že informácie obsiahnuté vo zvukovom podnete sa pri prechode rôznymi úrovňami sluchovej dráhy opakovane prekódujú. Počas tohto procesu neuróny jedného alebo druhého typu vyžarujú "svoje" vlastnosti stimulu, čo poskytuje pomerne špecifickú aktiváciu neurónov. vyššie úrovne.
V bežnom živote sa s čistými tónmi prakticky nestretávame. Zvuky okolo nás sa skladajú z rôznych frekvenčných zložiek, ktoré sa neustále a nezávisle menia. Ich amplitúda a trvanie sa tiež líšia; môžu prichádzať a odchádzať náhle alebo postupne, opakovať sa alebo byť jedinečné; ich zdroj sa môže nachádzať bližšie alebo ďalej od nás, pohybovať sa atď. Človek, aspoň s trénovaným sluchom, dokáže oceniť všetky tieto vlastnosti. Nervové procesy, ktoré sú základom takéhoto hodnotenia, boli identifikované najmä v sluchová kôra. Napríklad niektoré neuróny v primárnej sluchovej kôre reagujú len na začiatok zvukového podnetu, iné len na jeho koniec. Niektoré skupiny neurónov vzrušujú zvuky určitého trvania, iné opakujúce sa zvuky. Existujú aj bunky, ktoré sa aktivujú len tou či onou frekvenčnou alebo amplitúdovou moduláciou zvuku. Mnoho neurónov sa aktivuje v širokom rozsahu frekvencií, t.j. šum, v iných sa frekvenčno-prahové charakteristiky líšia v jednom alebo viacerých výrazných minimách. Väčšina kortikálnych buniek je excitovaná aferentami kontralaterálneho ucha, ale niektoré reagujú na ipsilaterálnu stimuláciu, zatiaľ čo iné reagujú len na bilaterálnu stimuláciu. Významná časť neurónov primárnej sluchovej kôry nie je aktivovaná žiadnymi experimentálnymi vplyvmi; možno sú vysoko špecifické a reagujú len na podnety, ktoré je príliš ťažké reprodukovať v laboratóriu.
Celkové reakcie buniek primárna sluchová kôra podobné tým, ktoré sú známe pre komplexné alebo superkomplexné neuróny vo zrakovej kôre. Je zrejmé, že sa podieľajú na rozpoznávaní sluchových vzorov, čo je proces, ktorý je veľmi dôležitý, napr. pre porozumenie reči. Aj v sluchovej kôre opíc sa našli bunky, ktoré reagujú najmä na zvuky spojené s vnútrodruhovou komunikáciou. Vlastnosti týchto neurónov však často závisia od niektorých neznámych parametrov a ich reakcie sa nepredvídateľne líšia.
Poškodenie spánkových lalokov mozgu, kde sa nachádza sluchová kôra, sťažuje porozumenie reči, priestorovej lokalizácii zdroja zvuku (pozri nižšie) a identifikácii jeho časových charakteristík. Takéto lézie však neovplyvňujú schopnosť rozlíšiť frekvenciu a silu zvuku. Centrálne spracovanie zvukových informácií sa podrobnejšie zaoberá v prácach.
Nedávne štúdie ukázali, že tonotopická organizácia kochley pretrváva na vyšších úrovniach sluchového systému, vrátane kôry. Prítomnosť takejto organizácie, t.j. usporiadaná distribúcia oblastí spojených s určitými zvukovými frekvenciami v primárnej sluchovej kôre bola predtým odmietnutá.
Ďalším výsledkom, na rozdiel od skorších predpokladov, bola skutočnosť, že sluchové neuróny vyšších úrovní nie sú charakterizované výraznými vrcholmi v charakteristikách frekvenčného prahu. V primárnej aferentácii sluchového nervu, ak je pokusné zviera v optimálnych podmienkach, sú veľmi odlišné.
Sluchová orientácia v priestore . Centrálny sluchový systém je veľmi dôležitý pre priestorovú orientáciu. Ako je známe z každodennej skúsenosti, binaurálne počúvanie smer k zdroju zvuku sa dá určiť celkom presne. Fyzický základ je smerovosť v tom, že zvyčajne je jedno ucho od neho ďalej ako druhé. Zvuk sa šíri konečnou rýchlosťou a dostáva sa do vzdialenejšieho ucha neskôr a s menej sila, a sluchové ústrojenstvo je schopné zistiť jeho rozdiel na dvoch ušiach už na úrovni 1 dB. Na obr. Obrázok 12.15 znázorňuje metódu výpočtu rozdielu v čase prenosu zvuku. Rozdiel vzdialeností Δ S = dsinα , kde (d je vzdialenosť medzi ušami aα - uhol, pod ktorým je zdroj zvuku umiestnený vzhľadom na objekt. Takže časové oneskorenie Δ t = ∆s /s, kde c je rýchlosť zvuku. Človek je schopný zachytiť oneskorenie len 3–10–5 s, čo zodpovedá odchýlke zdroja zvuku od strednej čiary asi o 3°. Za optimálnych podmienok možno rozlíšiť polovičný uhol.
Psychofyzikálne aj neurofyziologické experimenty ukázali, že smerové počúvanie je založené na rozdiel v čase vedenia a intenzite zvuku. Pri použití slúchadiel na stimuláciu každého ucha nezávisle, oneskorenie signálu alebo zníženie intenzity na jednej strane spôsobí, že zvuk sa lokalizuje v opačnom uchu. Oneskorenie možno kompenzovať zvýšením intenzity; v tomto prípade sa zdroj zvuku zdá byť umiestnený v hlave. Podobné výsledky sa získali v neurofyziologických experimentoch. V hornej olivovej, prvá úroveň

Ryža. 12.15. Výpočet rozdielu v čase, keď zvuk dosiahne pravé a ľavé ucho (pozri text)
sluchového ústrojenstva s obojstrannou aferentáciou existujú neuróny, ktoré sa správajú podobne vzhľadom na časové charakteristiky a intenzitu signálu. Vzrušenie v nich je maximálne, keď je zvuk v jednom uchu hlasnejší ako v druhom a predchádza mu. Druhý typ buniek je tu najaktívnejší, keď sa podnety zasahujúce do oboch uší určitým spôsobom líšia v čase a intenzite príchodu. To znamená, že prvý typ buniek maximálne reaguje na zvuk lokalizovaný pozdĺž osi jedného z uší a druhý na zvuk prichádzajúci pod určitým uhlom. V colliculi superior kvadrigeminy sa sluchové a zrakové aferentácie spájajú a vytvárajú trojrozmernú „mapu“ priestoru. IN sluchová kôra niektoré bunky sa tiež aktivujú len na veľmi špecifickom mieste vzhľadom na poslucháča zdroja zvuku. Pri jeho zničení trpí aj priestorová orientácia. Stále však nie je celkom jasné, ako si CNS poradí so stanovením časového rozdielu menej ako 10–4 s.
Rozdiely v čase a intenzite vedenia nestačia na pochopenie toho, či je zdroj zvuku pred alebo za, nad alebo pod hlavou. To si vyžaduje prídavné zariadenie - ušnicu. Jeho štruktúra „skresľuje“ signál takým spôsobom, v závislosti od miesta jeho zdroja, že ho možno lokalizovať. V technológii sa to dá využiť umiestnením mikrofónov do hlavy figuríny na miesto ušných bubienkov: stereo nahrávky získané s ich pomocou budú vynikajúcej kvality.
Sluch v hluku . Binaurálny sluch má aj inú, dôležitejšiu funkciu ako orientácia v priestore; pomáha analyzovať akustické informácie v prítomnosti vonkajšieho hluku. „Interaurálne“ rozdiely v intenzite a smere signálov využíva CNS na potlačenie hluku v pozadí a zvýraznenie užitočných zvukov (napríklad pri zameraní sa na správnu konverzáciu na preplnenom stretnutí). Tento selektívny proces filtrovania zvyšuje počuteľnosť približne o 10 dB. To sa nepočujúcim na jedno ucho nestáva, čo sa dá ľahko overiť zapchaním ucha. Preto je dôležité pri strate sluchu obnoviť binaurálny sluch napríklad pomocou načúvacích prístrojov.
Sluchová adaptácia . Sluchový systém, podobne ako iné zmyslové systémy, je schopný k adaptácii. Do tohto procesu sú zapojené periférne ucho aj centrálne neuróny. Adaptácia sa prejavuje prechodným zvýšením prahu sluchu. To je užitočné, pretože to znižuje prah pre rozlíšenie hlasitosti a tým uľahčuje diferenciáciu sluchových vnemov. V prispôsobenom uchu sú izofóny posunuté nahor a bližšie k sebe. Podrobnejšie informácie sú uvedené v dokumentoch.
Strata sluchu a hluchota majú veľmi významný vplyv na životy pacientov, a preto priťahujú veľkú pozornosť lekárov. Príčiny týchto porušení možno rozdeliť do troch kategórií.
1. Porušenie vedenia zvuku. Medzi ne patrí poškodenie stredného ucha. Napríklad pri zápale bubienko-kostný aparát neprenáša do vnútorného ucha normálne množstvo zvukovej energie. V dôsledku toho, aj keď je zdravý, sluch sa zhoršuje. Existujú mikrochirurgické spôsoby, ako takéto chyby sluchu efektívne odstrániť.
2. Porušenie vnímania zvuku. V tomto prípade sú vláskové bunky Cortiho orgánu poškodené, takže je narušené buď spracovanie signálu alebo uvoľňovanie neurotransmiterov. V dôsledku toho trpí prenos informácií z kochley do centrálneho nervového systému.
3. Retrokochleárne poruchy. Vnútorné a stredné ucho sú zdravé, ale postihnutá je buď centrálna časť primárnych aferentných vlákien alebo iné zložky sluchovej dráhy (napríklad pri nádore na mozgu).
Testovanie sluchu u pacientov je tzv audiometria. Na identifikáciu a lokalizáciu poškodenia načúvacieho prístroja bolo vyvinutých množstvo testov (podrobnejšie pozri).
Najdôležitejšie spomedzi nimi – prahová audiometria. Cez jedno slúchadlo sú pacientovi prezentované rôzne tóny. Lekár začína s jasne podprahovou intenzitou zvuku a postupne zvyšuje akustický tlak až do. pacient nebude hlásiť, že počuje zvuk. Tento akustický tlak je vynesený do grafu (obrázok 12.16) tzv audiogram.
Na štandardných audiografických formách je normálna úroveň sluchového prahu znázornená hrubou čiarou a označená ako „0 dB“. Na rozdiel od obr. 12.18 sú vyššie prahy zakreslené pod nulovou čiarou a charakterizujú stupeň straty sluchu - o koľko decibelov sú pod normálom. Zdôrazňujeme, že tu nehovoríme o decibeloch SPL, ale o strate sluchu toľkých dB. Ak si napríklad zapcháte obe uši prstami, zníži sa to asi o 20 dB (pri tomto experimente sa samozrejme treba snažiť nerobiť príliš veľa hluku samotnými prstami). Pomocou slúchadiel sa testuje vnímanie zvuku, keď je vedenie vzduchu. Kostné vedenie testované podobným spôsobom, ale namiesto slúchadiel sa používa ladička, priložená na testovacej strane na mastoidálny výbežok spánkovej kosti, takže vibrácie sa šíria priamo cez lebku. Porovnaním prahových kriviek pre kostné a vzdušné vedenie je možné rozlíšiť hluchotu spôsobenú poškodením stredného alebo vnútorného ucha.
Hluchota s poškodením stredného ucha je spôsobená porušením vedenia zvuku. Vnútorné ucho je však skvelé. Za takýchto podmienok sa strata sluchu zistí pri teste vedenia vzduchu (pozri obr. 12.16) a prah kostného vedenia je normálny, pretože zvuková energia pri testovaní dosiahne vláskové bunky a obíde stredné ucho.
Hluchota v patológii vnútorného ucha v dôsledku poškodenia vlasových buniek; stredné ucho je super. V tomto prípade je prah pre oba typy vedenia zvýšený, pretože v oboch prípadoch sa transformácia signálu uskutočňuje rovnakým receptorovým procesom. Retrokochleárne poruchy tiež zvyšujú oba prahové hodnoty.
Pomocou ladičky (zvyčajne pri 256 Hz) je možné veľmi ľahko rozlíšiť poruchy vedenia od poškodenia vnútorného ucha alebo retrokochleárnych patológií, keď je známe, ktoré ucho počuje najhoršie. (Weberov test). Noha oscilačnej ladičky je umiestnená na strednej línii lebky; ak je postihnuté vnútorné ucho, pacient hlási, že tón znie zo zdravej strany; ak priemer - s postihnutými.
Tento jav je ľahké vysvetliť v prípade patológie vnútorného ucha. Poškodené receptory menej vzrušujú sluchový nerv, preto v zdravom uchu znie tón hlasnejšie a vďaka tomuto rozdielu vzniká smerový vnem. V prípade lézie stredného ucha sa stretávame s tromi simultánnymi procesmi. Po prvé, zhoršenie vibračných vlastností kostného aparátu oslabuje prenos zvuku nielen zvonku dovnútra, ale aj v opačnom smere. Preto je vnútorné ucho vzrušené zvukom,
informácie v mozgu
Časť 2. Analýza vestibulárneho aparátu a zvuku
Anatómia vestibulárnej dráhy je mimoriadne zložitá (obr. 24). Aferentné vlákna z hrebeňov polkruhových kanálikov a makúl sacculus a utriculus sú vysielané v Scarpovom gangliu (vestibulárny) v blízkosti vonkajšieho zvukovodu, kde sa nachádzajú telá neurónov a následne po spojení s kochleárnymi vláknami vznikajú vestibulo-kochleárny nerv ísť do ipsilaterálny vestibulárny komplex nachádza sa vo ventrálnej časti medulla oblongata pod štvrtou mozgovou komorou. Komplex tvoria štyri dôležité jadrá: laterálne (Deitersovo jadro), mediálne, horné a zostupné. Existuje tiež veľa menších jadier spojených zložitým systémom aferentov a eferentov.
Tento komplex jadier je inervovaný zostupnými vláknami z cerebellum a retikulárnej formácie. Okrem toho každý komplex dostáva inervácia z kontralaterálneho komplexu . V niektorých prípadoch je táto kontralaterálna inervácia základom mechanizmu push-pull. Napríklad bunky hrebeňa polkruhového kanála tiež prijímajú informácie z hrebeňa kontralaterálneho kanála. K tomu všetkému ešte komplex prijíma informácie z očí a proprioceptívnych vlákien vzostupne po mieche. Vestibulárny komplex je teda mimoriadne dôležitým centrom pre integráciu informácií týkajúcich sa pohybu a orientácie. Ryža. 24 ukazuje, že okrem silných väzieb s cerebellum A okulomotorické jadrá, vestibulárny komplex posiela vlákna do mozgovej kôry. Verí sa, že končia v postcentrálny gyrus blízko dolného konca sulcus intraparietalis (intraparietálny sulcus). Záchvatom, ktoré sa zameriavajú na túto oblasť, zvyčajne predchádza aura (jedna zo zložiek epileptického záchvatu charakterizovaného zhoršeným vnímaním) charakterizovaná pocitmi točenia hlavy a dezorientácie.
vestibulárny aparát sleduje stacionárnu orientáciu hlavy v priestore (otolity) A zrýchlenie jeho pohybu (hrebene polkruhových kanálov). Toto všetko dopĺňajú informácie z mnohých somestetických receptorov v celom tele. Aby ste eliminovali tok informácií z týchto senzorov, musíte telo vložiť do vody alebo na orbitálnu stanicu. Za týchto podmienok všetka práca padá na oči a vestibulárny aparát; ak je teraz aj objekt zaslepený, zostane len informácia z membránovej predsiene.
Úlohu informácií z polkruhových kanálov možno názorne demonštrovať, ak subjekt sedí v rýchlo sa otáčajúcom kresle. V tomto prípade sa oči pohybujú v smere opačnom k rotácii v snahe fixovať nehybný objekt pohľadom a potom (keď sa stratí zo zorného poľa) rýchlo poskočia v smere rotácie, aby našli ďalší bod fixácie pohľadu. Podobne, keď sa rotácia náhle zastaví, oči pokračujú v smere predchádzajúcej rotácie a potom preskočia v opačnom smere. K tejto náhlej zmene dochádza v dôsledku skutočnosti, že hrebene polkruhových kanálikov sú ovplyvnené tokom endolymfy a obrátením smeru toku. Tieto charakteristické pohyby očí sa nazývajú nystagmus. Sú podmienené tri neurónové dráhy (Obr. 25):
Ø od polkruhových kanálikov k vestibulárnym jadrám,
Ø k vonkajším svalom očí.
Význam vestibulo-okulomotorický reflex možno názorne demonštrovať porovnaním videnia rotujúceho očného systému s videním, keď je hlava nehybná a prostredie sa otáča. Detaily rotujúceho prostredia sa stratia veľmi rýchlo: pri dvoch otáčkach za sekundu sa fixačný bod pohľadu rozmaže do rozmazania. Naproti tomu subjekt sediaci na otočnom kresle len mierne stráca zrakovú ostrosť pri rýchlosti rotácie asi 10 otáčok za sekundu.
Nakoniec stojí za to povedať pár slov kinetóza. Toto nepríjemný pocit sa vyskytuje najmä v dôsledku nezhody dotykového vstupu . V niektorých prípadoch sa tento nesúlad vyskytuje v samotnom vestibulárnom aparáte. Ak hlava stratí svoju normálnu orientáciu a otáča sa, signály z hrebeňov polkruhových kanálov už nekorelujú so signálmi z otolitov. Ďalším zdrojom kinetózy je nesúlad signálov z očí a z vestibulárneho aparátu. Ak na rozbúrenom mori v kabíne oči nehlásia žiadny relatívny pohyb medzi hlavou a stenami kabíny, zatiaľ čo vestibulárny aparát je naopak v strese, pozorujú sa príznaky „morskej choroby“. Za zmienku tiež stojí, že k nebezpečnej dezorientácii vedie aj nadmerná konzumácia alkoholu. Je to spôsobené skutočnosťou, že etanol mení špecifickú hustotu endolymfy, takže kupla teraz môže vnímať gravitáciu, a preto vysiela neobvyklé signály do centrálneho vestibulárneho systému.
Motorické reakcie očí na stimuláciu vestibulárneho aparátu (okulocefalický reflex, vestibulo-okulárny reflex) sú sprostredkované dráhami cez mozgový kmeň z vestibulárnych jadier medulla oblongata do jadier abducens a okohybných nervov. Normálne rotácia hlavy spôsobuje pohyb endolymfy v polkruhových kanáloch v opačnom smere ako rotácia. V tomto prípade v jednom labyrinte dochádza k toku endolymfy smerom k ampulke horizontálneho polkruhového kanála a v druhom labyrinte - v smere od ampulky kanála, zatiaľ čo podráždenie receptorov jedného kanála sa zvyšuje a podráždenie opačného kanála klesá, tzn dochádza k nerovnováhe impulzov prichádzajúcich do vestibulárnych jadier. Pri stimulácii vestibulárnych jadier na jednej strane sa informácia okamžite prenesie do kontralaterálneho jadra nervu abducens v mozgovom moste, odkiaľ impulzy cez mediálnu pozdĺžny zväzok dosahujú jadro okohybného nervu v medzimozgu na strane dráždivého vestibulárneho aparátu. Tým je zabezpečená synchrónna kontrakcia laterálneho priameho svalu oka oproti podráždenému labyrintu a stredného priameho svalu oka rovnakého mena, čo v konečnom dôsledku vedie k pomalému priateľskému vychýleniu očí v opačnom smere ako je smer hlavy. rotácia. Tento reflex vám umožňuje stabilizovať polohu očí a fixovať pohľad na nehybný predmet, a to aj napriek rotácii hlavy. U zdravého, bdelého človeka môže byť vplyvom mozgovej kôry na kmeňové štruktúry svojvoľne potlačené. u pacienta s jasnou mysľou sa integrita štruktúr zodpovedných za tento reflex určuje nasledovne. Pacient je požiadaný, aby uprel svoj pohľad na centrálne umiestnený predmet a rýchlo (dva cykly za sekundu) otočil hlavu pacienta buď jedným alebo druhým smerom. Ak je vestibulo-okulárny reflex zachovaný, pohyby očných bulbov sú plynulé, sú úmerné rýchlosti pohybov hlavy a smerujú opačným smerom. Na posúdenie tohto reflexu u pacienta v kóme sa používa bábkový očný test. Umožňuje vám určiť bezpečnosť funkcií stonky. Lekár fixuje hlavu pacienta rukami a otáča ju doľava a doprava, potom ju hodí späť a spustí ju dopredu; očné viečka pacienta by mali byť zdvihnuté (test je absolútne kontraindikovaný v prípadoch podozrenia na traumu krčnej chrbtice).
Test sa považuje za pozitívny, ak sa očné buľvy nedobrovoľne odchýlia v opačnom smere, ako je otočenie (fenomén "bábikových očí"). Pri intoxikácii a dysmetabolických poruchách s obojstrannými léziami mozgovej kôry je test „bábikové oko“ pozitívny (očné buľvy pacienta sa pohybujú v opačnom smere ako je smer rotácie hlavy). Pri léziách mozgového kmeňa chýba okulocefalický reflex, to znamená, že test je negatívny (očné gule sa pri otáčaní pohybujú súčasne s hlavou, ako keby boli zmrazené na mieste). Tento test je negatívny aj v prípade otravy niektorými lieky(napr. pri predávkovaní fenytoínom, tricyklickými antidepresívami, barbiturátmi, niekedy myorelaxanciami, diazepamom) však zostáva normálna veľkosť zreníc a ich reakcia na svetlo.
Kalorické testy sú tiež založené na reflexných mechanizmoch. Stimuláciu polkruhových kanálikov studenou vodou, ktorá sa nalieva do vonkajšieho ucha, sprevádza pomalé priateľské vychýlenie očných buliev smerom k podráždenému labyrintu. Studený kalorický test sa vykonáva nasledovne. Najprv sa musíte uistiť, že ušné bubienky v oboch ušiach nie sú poškodené. Pomocou malej injekčnej striekačky a krátkej tenkej hadičky z mäkkého plastu jemne vstreknite 0,2 – 1 ml do vonkajšieho zvukovodu ľadová voda. V tomto prípade zdravý bdelý človek bude mať nystagmus, ktorého pomalá zložka (pomalá odchýlka očných buľv) smeruje k podráždenému uchu a rýchla zložka je nasmerovaná opačným smerom (nystagmus, tradične určený rýchlou zložkou , smeruje opačným smerom). Po niekoľkých minútach postup zopakujte na opačnej strane. Tento test môže slúžiť ako expresná metóda na detekciu periférnej vestibulárnej hypofunkcie.
U pacienta v kóme, s neporušeným mozgovým kmeňom, tento test spôsobí tonicky koordinovanú odchýlku očných buliev smerom k ochladzovanému labyrintu, nedochádza však k rýchlym pohybom očí v opačnom smere (to znamená, že samotný nystagmus nie je pozorovaný) . Ak sú u pacienta v kóme poškodené štruktúry mozgového kmeňa, popísaný test nespôsobuje vôbec žiadne pohyby očných bulbov (nedochádza k tonickej odchýlke očných bulbov).
vestibulárna ataxia
Vestibulárna ataxia sa zisťuje pomocou Rombergovho testu a štúdia chôdze pacienta (ponúkajú mu, aby kráčal v priamej línii s otvorenými očami a potom so zatvorenými očami). Pri jednostrannej periférnej vestibulárnej patológii sa pozoruje nestabilita pri státí a chôdzi v priamom smere s odchýlkou smerom k postihnutému labyrintu. Vestibulárna ataxia je charakterizovaná zmenou závažnosti ataxie s náhlymi zmenami polohy hlavy a otočením pohľadu. Uskutočňuje sa aj indexový test: subjekt je požiadaný, aby zdvihol ruku nad hlavu a potom ju spustil, pričom sa snaží dostať ukazovák do ukazováka lekára. Prst lekára sa môže pohybovať rôznymi smermi.
Najprv pacient vykoná test s otvorenými očami, potom je požiadaný, aby test vykonal so zatvorenými očami. Pacient s vestibulárnou ataxiou míňa oboma rukami smerom k pomalej zložke nystagmu.
Statické a statokinetické reflexy. Rovnováha je udržiavaná reflexne, bez zásadnej účasti vedomia na tomto. Existujú statické a statokinetické reflexy. S oboma sú spojené vestibulárne receptory a somatosenzorická aferentácia, najmä z cervikálnych proprioceptorov. Statické reflexy zabezpečujú primeranú vzájomnú polohu končatín, ako aj stabilnú orientáciu tela v priestore, t.j. posturálne reflexy. Vestibulárna aferentácia pochádza v tomto prípade z otolitických orgánov. Statický reflex, ktorý je ľahko pozorovateľný u mačky vďaka vertikálnemu tvaru jej zrenice, je kompenzačná rotácia očnej gule, keď sa hlava otáča okolo dlhej osi tela (napríklad so sklopeným ľavým uchom). Žiaci zároveň po celý čas zachovávajú polohu veľmi blízko vertikále. Tento reflex sa pozoruje aj u ľudí. Statokinetické reflexy sú reakcie na motorické podnety, ktoré sa samy prejavujú v pohyboch. Sú spôsobené excitáciou receptorov polkruhových kanálikov a otolitických orgánov; Príkladmi sú rotácia mačacieho tela pri páde, ktorá zabezpečí, aby dopadla na všetky štyri nohy, alebo pohyb človeka, ktorý znovu nadobudne rovnováhu po tom, čo zakopne.
Jedným zo statokinetických reflexov je vestibulárny nystagmus. Ako bolo uvedené vyššie, vestibulárny systém spôsobuje rôzne pohyby očí; nystagmus, ako ich špeciálna forma, sa pozoruje na začiatku rotácie, ktorá je intenzívnejšia ako bežné krátke otáčky hlavy. Oči sa v tomto prípade otáčajú proti smeru rotácie, aby zachovali pôvodný obraz na sietnici, avšak nedosiahnuc krajnú možnú polohu, náhle „poskočia“ v smere rotácie a objaví sa ďalší úsek priestoru. zorné pole. Potom nasleduje ich pomalý spätný pohyb.
Pomalú fázu nystagmu spúšťa vestibulárny systém a rýchle „skákanie“ pohľadu spúšťa prepontínna časť retikulárnej formácie.
Pri rotácii tela okolo zvislej osi sú podráždené takmer len horizontálne polkruhové kanáliky, t.j. odchýlka ich kupúl spôsobuje horizontálny nystagmus. Smer oboch jeho zložiek (rýchla a pomalá) závisí od smeru rotácie a tým aj od smeru deformácie kupule. Ak sa telo otáča okolo horizontálnej osi (napríklad pri prechode cez uši alebo sagitálne cez čelo), dochádza k stimulácii zvislých polkruhových kanálikov a vzniku vertikálneho alebo rotačného nystagmu. Smer nystagmu je zvyčajne určený jeho rýchlou fázou, t.j. pri „pravom nystagme“ pohľad „skočí“ doprava.
Pri pasívnej rotácii tela vedú k výskytu nystagmu dva faktory: stimulácia vestibulárneho aparátu a pohyb zorného poľa vzhľadom na osobu. Optokinetický (spôsobený zrakovou aferentáciou) a vestibulárny nystagmus pôsobia synergicky.
Diagnostická hodnota nystagmu. Nystagmus sa používa na klinike na testovanie vestibulárnej funkcie. Subjekt sedí v špeciálnom kresle, ktoré sa dlho otáča konštantnou rýchlosťou a potom sa náhle zastaví. Zarážka spôsobí, že sa kopula odchýli v opačnom smere, než v akom sa odchýlila na začiatku pohybu; výsledkom je nystagmus. Jeho smer môže byť určený registráciou deformácie kopule; musí byť proti smeru predchádzajúceho pohybu. Záznam očných pohybov sa podobá záznamu získanému v prípade optokinetického nystagmu. Nazýva sa to nystagmogram.
Po testovaní na postrotačný nystagmus je dôležité vylúčiť možnosť fixácie pohľadu v jednom bode, keďže zraková aferentácia dominuje nad vestibulárnou aferentáciou pri okulomotorických reakciách a za určitých podmienok môže nystagmus potlačiť. Preto je objekt nasadený na okuliare Frenzel s vysoko vypuklými šošovkami a vstavaným zdrojom svetla. Robia ho „krátkozrakým“ a neschopným fixovať svoj pohľad, pričom lekárovi umožňujú ľahko pozorovať pohyby očí. Takéto okuliare sa vyžadujú aj pri teste spontánneho nystagmu, prvom, najjednoduchšom a najdôležitejšom postupe pri klinickom vyšetrení vestibulárnej funkcie.
Ďalším klinickým spôsobom na spustenie vestibulárneho nystagmu je tepelná stimulácia horizontálnych polkruhových kanálov. Jeho výhodou je možnosť otestovať každú stranu tela zvlášť. Hlava sediaceho je naklonená dozadu približne o 60° (u ležiaceho na chrbte je zdvihnutá o 30°), takže horizontálny polkruhový kanál je v striktne vertikálnom smere. Potom sa vonkajší zvukovod umyje studenou alebo teplou vodou. Vonkajší okraj polkruhového kanálika je veľmi blízko k nemu, takže sa okamžite ochladzuje alebo zahrieva. Podľa Baraniho teórie hustota endolymfy pri zahrievaní klesá; následne jeho vyhrievaná časť stúpa a vytvára tlakový rozdiel na oboch stranách kuply; výsledná deformácia spôsobuje nystagmus. Na základe svojej povahy sa tento typ nystagmu nazýva kalorický. Pri zahrievaní smeruje do miesta tepelného dopadu, pri ochladzovaní opačným smerom. U ľudí trpiacich vestibulárnymi poruchami sa nystagmus kvalitatívne a kvantitatívne líši od normálu. Podrobnosti o jeho testovaní sú uvedené v článku. Treba poznamenať, že kalorický nystagmus sa môže vyskytnúť v kozmickej lodi v podmienkach beztiaže, keď sú rozdiely v hustote endolymfy nevýznamné. V dôsledku toho sa na jeho spustení podieľa ešte aspoň jeden, zatiaľ neznámy mechanizmus, napríklad priamy tepelný účinok na vestibulárny orgán.
Funkciu otolitického aparátu možno testovať pozorovaním okulomotorických reakcií pri záklone hlavy alebo pri pohybe pacienta tam a späť na špeciálnej plošine.




