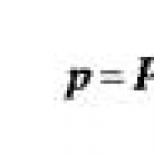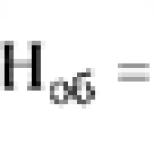Bakterilerde spesifik transdüksiyon. Bakteriyofajlar, yapısal özellikler ve pratik uygulama. Anaerobik bakterilerin fizyolojisinin özellikleri
Spesifik transdüksiyon 1956'da M. Morse ve eşleri E. ve J. Lederberg tarafından keşfedildi. Karakteristik özellik spesifik transdüksiyon, her transdüksiyon fajının bakteri kromozomunun sadece belirli, çok sınırlı bir bölgesini iletmesidir. Genelleştirilmiş transdüksiyonda faj, bakterilerin genetik materyalinin "pasif" bir taşıyıcısı olarak hareket ederse ve dönüştürülmüş bakterilerdeki genetik rekombinasyon, rekombinasyon sürecinin genel modellerine göre gerçekleşirse, o zaman spesifik transdüksiyon durumunda, faj sadece genetik materyali aktarır, ancak aynı zamanda bakteri kromozomuna dahil edilmesini sağlar. Spesifik transdüksiyonun en iyi bilinen örneği, E. coli bakteri hücrelerini enfekte edebilen ve DNA'sının bakteri genomuna entegrasyonunu sağlayan λ faj tarafından gerçekleştirilen transdüksiyondur. Bakterilerin lizojenizasyonu sırasında, bölgeye özgü rekombinasyonun (DNA iplikçiklerinin kırılması ve çapraz birleşmesinin) bir sonucu olarak ılıman faj λ, kromozomlarına yalnızca tek bir yerde entegre edilir: biyo ve gal lokusları arasındaki alanda. Bu alana attλ denir. Profajın indüksiyonu sırasında kromozomdan profajın eksizyonu (eksizyonu) ayrıca bölgeye özgü rekombinasyon mekanizmasına göre gerçekleştirilir. Siteye özel yeniden birleştirme hatasız değil, doğru bir şekilde gerçekleşir. Profaj eksizyon sırasında milyon olay başına yaklaşık bir kez, rekombinasyon attλ bölgesinde değil, gal veya bio bölgelerini yakalar. Bunun, profajın dağılması sırasında ilmeklerin “yanlış” oluşumundan kaynaklandığına inanılmaktadır. Sonuç olarak, bakteri genomunun profajın bitişiğindeki bölgesi kromozomdan ayrılır ve serbest faj genomunun bir parçası olur. Döngüdeki konumuna karşılık gelen profaj genomunun bölgesi bakteri kromozomunda kalır. Böylece, profaj ile bakteri kromozomu arasında genetik bir değişim gerçekleşir. Faj genomuna entegre olan bakteriyel genetik materyal, faj genetik materyalinin 1/3'üne kadar yerini alabilir. Bir kısmı bakteriyel DNA ile değiştirilen faj DNA'sının paketlenmesinden sonra, faj kafasında kusurlu faj parçacıkları oluşturulur. Faj, kafa hacminin sınırlı olması ve genomuna bir bakteri DNA parçası dahil edildiğinde, faj genomunun bir kısmının bakteri kromozomunda kalması nedeniyle kusurludur. Kusur önemsiz ise, protein kaplaması bozulmadan kaldığı ve hücreler üzerinde adsorpsiyon sağladığı için faj canlı kalır. Böyle kusurlu bir faj, diğer hücreleri enfekte edebilir, ancak üremeden sorumlu genler olmadığı için üreme enfeksiyonuna neden olamaz. Böyle kusurlu bir fajda, dairesel bir forma dönüşmesini sağlayan DNA yapışkan uçları korunursa, kusurlu fajın DNA'sı, bir bakteri DNA parçası ile birlikte, alıcı bakterilerin DNA'sına entegre olabilir ve onların lizojenleşmesine neden olabilir. gal lokusunun genlerini içeren kusurlu parçacıklar oluşur. Bu tür kusurlu parçacıklar, λdgal (faj λ, kusurlu, gal) olarak adlandırılır. Faj λ genomu, biyotin sentezinden sorumlu geni içeriyorsa, λdbio olur. Bu nedenle, alıcı hücreler, donör bakterilerin kusurlu partiküller içeren faj λ ile enfeksiyonundan sonra elde edilen bir fagolizat ile bio– veya gal– ile tedavi edilirse, 10–5–10–6 sıklıkta transdüktanlar bio+ veya gal+ oluşur. E. coli'de spesifik transdüksiyon sadece λ faj tarafından değil, aynı zamanda lambdoid fajlar olarak adlandırılan ve φ80, 434, 82, vb. dahil ilgili fajlar tarafından da gerçekleştirilir. Özellikle, φ80 faj, kodlayan genlerin yakınında kromozoma dahil edilir. triptofanın sentezinden sorumlu enzimlerin oluşumu. Bu nedenle φ80 faj trp genlerinin transferi için uygundur. S. typhimurium'un P22 fajının, genel transdüksiyona ek olarak spesifik transdüksiyonu da gerçekleştirebildiği bulunmuştur. Litik gelişim döngüsü sırasında, bakteriyofaj P22 genel transdüksiyon gerçekleştirebilirken, lizojenizasyon sırasında spesifik transdüksiyon gerçekleştirebilir. P22 faj DNA'sı, prolin sentezinden sorumlu genlerin yanında kromozomun bir bölgesine entegre edilmiştir. Profaj entegrasyonu, spesifik dönüştürücü parçacıkların oluşumunu önemli ölçüde uyarır. Bu nedenle, spesifik transdüksiyon, donör bakterilerin ön lizojenizasyonunu ve ardından hücrelerden profajın indüklenmesini gerektirir. Ortaya çıkan kusurlu dönüştürücü faj partikülleri, alıcı suşun hücrelerini enfekte eder, bunlar lizojenize edilir ve profaj, donörün bakteriyel genomunun bir kısmı ile alıcının kromozomuna eklenir. Transdüksiyon aşağıdaki yönlerde kullanılabilir: donör kromozomunun plazmitlerini ve kısa parçalarını transdükte edin; belirli bir genotipin suşlarının, özellikle izogenik suşların inşası için. Burada, aktarılan parçaların küçük boyutu, konjugasyona göre transdüksiyon avantajı sağlar. Genelleştirilmiş transdüksiyon kullanılarak oluşturulan izogenik suşlar, yalnızca dönüştürücü faj tarafından taşınan kromozom bölgesinde farklılık gösterir; bakteri genlerinin doğru haritalanması, sıranın ve operonlardaki konumlarının belirlenmesi için ve iyi yapı bir tamamlama testi kullanılarak gerçekleştirilen bireysel genetik belirleyiciler. Belirli bir ürün grubunun sentezinin birkaç genin işleyişini gerektirdiği bilinmektedir. Bazı enzimlerin sentezinin a ve b genlerinin ürünleri tarafından belirlendiğini varsayalım. Enzim sentezleyemeyen iki fenotipik olarak özdeş mutant olsun, ancak bunların genetik olarak aynı mı yoksa farklı mı oldukları bilinmiyor. Genotipi belirlemek için transdüksiyon gerçekleştirilir, yani faj bir popülasyonun hücreleri üzerinde yayılır ve ardından ikinci popülasyonun hücreleri fagolizat ile enfekte edilir. Seçici bir ortam üzerinde aşılama üzerine hem büyük gerçek transdüktanlar kolonileri hem de küçük abortif transdüktanlar kolonileri oluşursa, mutasyonların farklı genlerde lokalize olduğu sonucuna varılır.
Ders kitabı yedi bölümden oluşmaktadır. Birinci Bölüm - "Genel Mikrobiyoloji" - bakterilerin morfolojisi ve fizyolojisi hakkında bilgiler içerir. İkinci bölüm bakterilerin genetiğine ayrılmıştır. Üçüncü bölüm - "Biyosferin mikroflorası" - çevrenin mikroflorasını, doğadaki maddelerin döngüsündeki rolünü ve ayrıca insan mikroflorasını ve önemini dikkate alır. Dördüncü bölüm - "Enfeksiyon doktrini" - mikroorganizmaların patojenik özelliklerine, bulaşıcı süreçteki rollerine ayrılmıştır ve ayrıca antibiyotikler ve etki mekanizmaları hakkında bilgi içerir. Beşinci bölüm - "Bağışıklık Doktrini" - bağışıklık hakkında modern fikirleri içerir. Altıncı bölüm - "Virüsler ve sebep oldukları hastalıklar" - virüslerin temel biyolojik özellikleri ve sebep oldukları hastalıklar hakkında bilgi vermektedir. Yedinci bölüm - "Özel tıbbi mikrobiyoloji" - birçok patojenin morfolojisi, fizyolojisi, patojenik özellikleri hakkında bilgi içerir. bulaşıcı hastalıklar, modern teşhis yöntemlerinin yanı sıra spesifik önleme ve tedavi.
Ders kitabı öğrenciler, yüksek lisans öğrencileri ve yüksek tıp öğretmenleri için tasarlanmıştır. Eğitim Kurumları, üniversiteler, tüm uzmanlık alanlarından mikrobiyologlar ve uygulayıcılar.
5. baskı, gözden geçirilmiş ve genişletilmiş
Kitap:
| <<< Назад
|
İleri >>> |
Bu durumda, transdüksiyon fajlarının her zaman sadece belirli genleri, yani attL'nin solunda veya attR'nin sağında lizojenik hücrenin kromozomunda bulunan genleri taşıması spesifik olmayandan farklıdır. Spesifik transdüksiyon her zaman ılıman fajın konak hücrenin kromozomuna entegrasyonu ile ilişkilidir. Kromozomdan ayrılırken (dışlama) profaj, geni sol veya sağ kanattan, örneğin gal veya bio'dan yakalayabilir. Ancak bu durumda, toplam uzunluğunun değişmemesi için DNA'sının aynı boyutunu karşı uçtan kaybetmesi gerekir (aksi takdirde faj kafasına paketlenemez). Bu nedenle, bu dışlama biçimiyle kusurlu fajlar oluşur: ?dgal veya ?dbio.
spesifik transdüksiyon E. koli sadece lambda fajını değil, aynı zamanda ilgili lambdoid ve diğer fajları da gerçekleştirir. AttB bölgelerinin kromozom üzerindeki konumuna bağlı olarak, dışlandıklarında, çeşitli profaj bağlantılı bakteri genlerini açabilir ve onları diğer hücrelere aktarabilirler. Genoma dahil edilen materyal, fajın genetik materyalinin 1/3'üne kadar yerini alabilir.
Alıcı hücrenin enfeksiyonu durumunda dönüştürücü faj, kromozomuna entegre olur ve ona yeni bir gen (yeni özellik) sokar, sadece lizojenizasyona değil, aynı zamanda lizojenik dönüşüme de aracılık eder.
Bu nedenle, spesifik olmayan transdüksiyon sırasında faj sadece genetik materyalin pasif bir taşıyıcısıysa, o zaman spesifik transdüksiyon sırasında faj bu materyali genomuna dahil eder ve onu, lizojenleştirici bakterileri alıcıya aktarır. Bununla birlikte, ılıman faj genomu, hücrede bulunmayan ancak temel proteinlerin sentezinden sorumlu olan kendi genlerini içeriyorsa, lizojenik dönüşüm de meydana gelebilir. Örneğin, yalnızca difteri patojenleri, kromozomunda toksin operonunu taşıyan ılımlı bir profajın entegre edildiği eksotoksin üretme yeteneğine sahiptir. Difteri toksininin sentezinden sorumludur. Başka bir deyişle, ılıman toksik faj, toksik olmayan difteri basilinin lizojenik dönüşümüne neden olur.
Agar tabakası yöntemi aşağıdaki gibidir. İlk olarak, tabağa bir besin agar tabakası dökülür. Katılaşmadan sonra, önce bir damla konsantre bakteri süspansiyonu ve belirli bir hacimde faj süspansiyonunun eklendiği bu katmana, eritilmiş ve 45 °C'ye soğutulmuş 2 ml %0.7 agar eklenir. Üst tabaka sertleştikten sonra fincan bir termostata yerleştirilir. Bakteriler, agarın yumuşak tabakası içinde çoğalarak faj kolonilerinin steril noktalar şeklinde açıkça görülebildiği sürekli bir opak arka plan oluşturur (Şekil 84, 2). Her koloni, bir ana faj virionunun çoğaltılmasıyla oluşturulur. Bu yöntemin kullanılması şunları sağlar: a) belirli bir materyaldeki canlı faj viryonlarının sayısını doğru bir şekilde belirlemek için kolonileri sayarak;
b) tarafından özellikler(boyut, şeffaflık, vb.) fajlardaki kalıtsal değişkenliği incelemek için.
Bakteriler üzerindeki etki spektrumuna göre fajlar ikiye ayrılır: çok değerli(liz ile ilgili bakteriler, örneğin, çok değerlikli Salmonella faj, hemen hemen tüm Salmonella'ları lize eder), monofajlar(sadece bir türün bakterilerini lize ederler, örneğin, Vi-I faj sadece patojenleri lize eder) Tifo) ve türe özgü Bir tür içindeki bakteri varyantlarını seçici olarak parçalayan fajlar. Bu tür fajların yardımıyla, bir tür içindeki bakterilerin en ince farklılaşması, faj varyantlarına bölünmeleriyle birlikte gerçekleştirilir. Örneğin, bir dizi Vi-II faj kullanılarak, tifoya neden olan ajan 100'den fazla faj varyantına bölünür. Bakterilerin fajlara duyarlılığı, karşılık gelen reseptörlerin mevcudiyeti ile ilişkili nispeten kararlı bir özellik olduğundan, faj tiplendirmesi büyük tanısal ve epidemiyolojik öneme sahiptir.
Pirinç. 84. Test materyalinde bakteriyofajların tespiti:
1 - nokta testi; 2 - Grazia'ya göre titrasyon
| <<< Назад
|
İleri >>> |
Genel transdüksiyonda, konak hücrenin DNA'sının segmentlerini içeren faj partikülleri, bir bakteri hücresinden diğerine nispeten uzun genomik DNA uzantıları aktarır. Dönüştürücü faj parçacıkları, hücrenin DNA'sı etkin bir şekilde bozunduğunda ve parçalandığında, belirli bulaşıcı süreçler sırasında oluşur.
Hücresel DNA, yaklaşık olarak faj genomunun büyüklüğü, tesadüfen olgun bakteriyofaj parçacıklarına paketlenir. Dönüştürücü fajları içerenler de dahil olmak üzere bir faj partikülü popülasyonu ile bakteri hücrelerinin sonraki enfeksiyonunun bir sonucu olarak, ikincisinin yardımıyla donör hücrelerin DNA'sı bu enfekte olmuş hücrelere aktarılır. Verici DNA'nın sokulan parçaları ile alıcı hücrenin DNA'sı arasındaki rekombinasyon, alıcı hücrenin genotipinde bir değişikliğe yol açar.
Her dönüştürücü faj parçacığı genellikle orijinal donör kromozomunun yalnızca bir rastgele parçasını içerir. Verici genomun herhangi bir parçasını böyle bir partiküle dahil etme olasılığı yaklaşık olarak aynıdır. Bununla birlikte, dönüştürülmüş DNA segmentlerinin oldukça büyük boyutu nedeniyle (belirli bakteriyofajlar için yaklaşık 100 kb veya tüm E. coli kromozomunun yüzde 2.5'i kadardır), alıcı hücre genellikle bir transdüksiyon olayında bütün bir gen grubunu edinir. Sonuç olarak, donör kromozom üzerinde birbirine yakın olan genler yüksek bir frekansla birlikte transdüksiyona tabi tutulurken, birbirinden uzak olan genler bağımsız olarak transdüksiyona tabi tutulur. Gen birlikte transdüksiyon sıklığının belirlenmesi, yakından bağlantılı genler arasındaki göreceli mesafelerin tahminine izin vererek genetik haritaların düzeltilmesine yardımcı olur. 3 Spesifik (sınırlı) transdüksiyon
Spesifik ikinci tipin transdüksiyonu, virüs genomunun enfekte hücrenin spesifik bir kromozomal DNA lokusuna dahil edilmesinin bir sonucu olarak bulaşıcı döngüsü kesintiye uğrayan ılıman bakteriyofajların karakteristiğidir. Bu tür entegre faj genomlarını içeren bakterilere denir. lizojenik. Viral genomları kendi kromozomlarının kalıtsal unsurları olarak taşırlar. Lizojenik bir hücrede viral ve hücresel genomlar tek bir birim olarak çoğalır ve karşılıklı olarak uyumludur. Faj genomunun konak hücrenin genomu ile entegrasyonu, fajın hücre ölümünü indükleme ve enfeksiyöz yavrular üretme yeteneğinden mahrum kalır. Bu nedenle bakteriyofaj
aksine lizogenez yeteneğine sahip öldürücü faj, adını aldım ılıman.
Belirli şartlar altında - indüksiyon- lizojenik durum kesintiye uğrar ve viral genom, konakçı kromozomdan çıkarılır. Birçok viral partikül oluşturmak için çoğalır ve hücreyi öldürür. Genellikle viral genomun kesilmesi çok kesindir ve ortaya çıkan faj, orijinaline tam olarak karşılık gelen bir viral genom içerir.
Bazen faj genomu yanlış kesilir ve kromozomal genler yavru faj parçacıklarına dahil edilir, bitişik entegre viral genoma. Bu genler, bazı viral genlerin yerine açılır. Bir sonraki enfeksiyon döngüsü sırasında, donör hücrenin genleri, alıcı hücrelere faj genleriyle birlikte geçer. Dönüştürücü fajın DNA'sının alıcı genoma dahil edilmesinden sonra hücre, faj genomu ile birlikte önceki faj konağının genetik bilgisini edinir.
Bu nedenle, spesifik transdüksiyonda faj, bir hücreden diğerine gen transferi için bir vektör görevi görür. Bu mekanizma kullanılarak, yalnızca viral genomun entegrasyon bölgesine yakından bağlı olan konakçı hücrenin kromozomal genleri dönüştürülür.
Farklı ılıman fajlar farklı kromozom bölgelerine girdiğinden, bunların yanlış kesilmesi, farklı kromozomal genleri dönüştüren fajlarla sonuçlanacaktır. yani fajlar lambda galaktoz metabolizmasından sorumlu genleri veya biotin sentezini kontrol eden genleri ve f80 fajlarını - triptofan biyosentezi için enzimleri kodlayan farklı sayıda genleri iletir.
Faj genomu, aşağıdakilerin sağlanması koşuluyla spesifik transdüksiyon yeteneğine sahiptir:
1 Dönüştürülmek için kovalent olarak bağlı viral olmayan bir DNA segmenti edinmelidir. DNA'nın bu bölümü genellikle hücresel kökenlidir, ancak prensipte herhangi bir kaynaktan olabilir. Olduğu sürece viral genomun herhangi bir yerine dahil edilebilir.
enfekte olmuş bir konak hücrede viral DNA'nın replikasyonunu veya olgun faj partikülleri halinde paketlenme yeteneğini etkilemez.
2 Faj genomu, alıcı hücrenin enfeksiyonu meydana geldikten sonra çoğalabilmelidir, yani. Viral DNA, replikasyonun orijinini (OP) ve replikasyonun gerçekleşmesi için gerekli genleri korumalıdır.
3 Yapısal faj proteinlerini kodlayan faj genleri fonksiyonel olarak aktif olmalıdır.
Spesifik transdüksiyon moleküler genetikte yaygın olarak kullanılmaktadır. Bu fenomenin böyle bir uygulamasının bir örneğini düşünün. Beta-galaktosidaz enziminin sentezini kodlayan Escherichia coli geni 3600 bp içerir. ve bu mikroorganizmanın genomunun binde biridir. Beta-galaktosidaz sentezini kodlayan bir bakteri hücresinin DNA parçası, dönüştürücü bir bakteriyofaj lambda genomuna yerleştirilirse, bunun on beşte birini kaplar, yani lambda fajının DNA'sı beta-galaktosidaz geninde zenginleşir. Escherichia coli'nin DNA'sından 100 kat daha fazla.
Transdüksiyon, bir bakteriyofaj tarafından bir bakteri hücresinden diğerine genlerin aktarılmasıdır. Bu fenomen ilk olarak 1952'de N. Zinder ve J. Lederberg tarafından kurulmuştur. Fareler için patojen olan Salmonella typhimurium bakterileri üzerinde çalışmalar yaptılar. Bu bakterilerin iki suşu seçildi: triptofan (T ~) sentezleyemeyen oksotropik suşu 22A ve triptofanı (T 1 ") sentezleyebilen suşu 2A. Bu suşlar, altta ayrılmış U şeklinde bir tüpe ekildi. bir bakteri filtresi ile (Şekil 24 22A suşu (T~), tüpün bir dizine ve 2A suşu (T1 ") diğerine aşılandı. Belirli bir kuluçka döneminden sonra, 22A suşu bakterileri, minimal bir besin ortamına ekildiğinde az sayıda koloni verdi (dönüştürülmüş hücrelerin görülme sıklığı N0~5 idi). Bu, bazı hücrelerin triptofan sentezleme yeteneğini kazandığını gösterdi. Bakteriler bu özelliği nasıl elde edebilir? Araştırma
Pirinç. 24. Transdüksiyon deneyi şeması
suş 22A'nın P-22 faj için lizojenik olduğunu gösterdi. Bu
Faj, lizojenik kültürden serbest bırakıldı,
filtre ve parçalanmış soy 2A. Genetiğin bir kısmını ekleyerek
2A suşundan, Bakteriyel faj geri geldi ve bu genetik materyali 22A suşuna aktardı. 22A'da gerinim
soy 2A'nın edinilmiş spesifik kalıtsal özellikleri,
bu durumda triptofan sentezleme özelliği. Diğer özellikler, yetenek dahil olmak üzere benzer bir şekilde dönüştürülebilir.
fermantasyon, antibiyotik direnci vb.
İletim fenomeni, Escherichia coli ve actinomycetes'te de kurulmuştur. Kural olarak, bir gen, nadiren iki ve çok nadiren üç bağlantılı gen dönüştürülür. Genetik materyalin transferi sırasında faj DNA molekülünün bir kısmı değiştirilir. Faj kendi parçasını kaybeder ve kusurlu hale gelir. Genetik materyalin alıcı bakterinin kromozomuna dahil edilmesi, çaprazlama gibi bir mekanizma ile gerçekleştirilir. Alıcının kromozomunun homolog bölgeleri ile faj tarafından verilen materyal arasında kalıtsal materyal alışverişi vardır.
Üç tür transdüksiyon vardır: genel veya spesifik olmayan, spesifik ve abortif. Faj partiküllerinin montajı sırasında spesifik olmayan transdüksiyon sırasında, etkilenen bakterinin DNA fragmanlarından herhangi biri, faj DNA'sı ile birlikte başlarına dahil edilebilir. Sonuç olarak, donör bakterinin çeşitli genleri alıcı hücrelere aktarılabilir. Spesifik olmayan transdüksiyon, Escherichia, Shigella ve Salmonella'daki P-1 ve P-22 fajları tarafından gerçekleştirilebilir. Spesifik transdüksiyon ile, profaj bakteri kromozomu üzerinde belirli bir yere dahil edilir ve profajın yanındaki donör hücrenin kromozomunda bulunan belirli genleri transdükte eder. Örneğin, profaj durumundaki "k (lambda) fajı, Escherichia coli'nin kromozomunda her zaman aynı yere dahil edilir ve galaktozu fermente etme yeteneğini belirleyen bir lokus dönüştürür. Profajlar konakçı DNA'dan ayrıldığında, profaj'a bitişik bakteriyel genler, onunla birlikte bileşim kromozomlarından ayrılır ve profaj genlerinin bir kısmı bileşiminde kalır.Toplam transdüksiyon sıklığı 1 milyonda 1'den 100 milyonda 1'e kadardır.Spesifik transdüksiyon daha sık meydana gelir.
Alıcının hücresine aktarılan donörün kromozomunun bir parçasının her zaman alıcının kromozomuna dahil edilmediği, ancak hücrenin sitoplazmasında korunabileceği tespit edilmiştir. Bakteriler bölündüğünde, yavru hücrelerden sadece birine girer. Bu duruma abortif transdüksiyon denir.
Spesifik transdüksiyon
Bu durumda, transdüksiyon fajlarının her zaman sadece belirli genleri, yani attL'nin solunda veya attR'nin sağında lizojenik hücrenin kromozomunda bulunan genleri taşıması spesifik olmayandan farklıdır. Spesifik transdüksiyon her zaman ılıman fajın konak hücrenin kromozomuna entegrasyonu ile ilişkilidir. Kromozomdan ayrılırken (dışlama) profaj, geni sol veya sağ kanattan, örneğin gal veya bio'dan yakalayabilir. Ancak bu durumda, toplam uzunluğunun değişmemesi için DNA'sının aynı boyutunu karşı uçtan kaybetmesi gerekir (aksi takdirde faj kafasına paketlenemez). Bu nedenle, bu dışlama biçimiyle, arr.
spesifik transdüksiyon E. koli sadece lambda fajını değil, aynı zamanda ilgili lambdoid ve diğer fajları da gerçekleştirir. AttB bölgelerinin kromozom üzerindeki konumuna bağlı olarak, dışlandıklarında, çeşitli profaj bağlantılı bakteri genlerini açabilir ve onları diğer hücrelere aktarabilirler. Genoma dahil edilen materyal, fajın genetik materyalinin 1/3'üne kadar yerini alabilir.
Alıcı hücrenin enfeksiyonu durumunda dönüştürücü faj, kromozomuna entegre olur ve ona yeni bir gen (yeni özellik) sokar, sadece lizojenizasyona değil, aynı zamanda lizojenik dönüşüme de aracılık eder.
Bu nedenle, spesifik olmayan transdüksiyon sırasında faj sadece genetik materyalin pasif bir taşıyıcısıysa, o zaman spesifik transdüksiyon sırasında faj bu materyali genomuna dahil eder ve onu, lizojenleştirici bakterileri alıcıya aktarır. Bununla birlikte, ılıman faj genomu, hücrede bulunmayan ancak temel proteinlerin sentezinden sorumlu olan kendi genlerini içeriyorsa, lizojenik dönüşüm de meydana gelebilir. Örneğin, yalnızca difteri patojenleri, kromozomunda toksin operonunu taşıyan ılımlı bir profajın entegre edildiği eksotoksin üretme yeteneğine sahiptir. Difteri toksininin sentezinden sorumludur. Başka bir deyişle, ılıman toksik faj, toksik olmayan difteri basilinin lizojenik dönüşümüne neden olur.
Agar tabakası yöntemi aşağıdaki gibidir. İlk olarak, tabağa bir besin agar tabakası dökülür. Katılaşmadan sonra, önce bir damla konsantre bakteri süspansiyonu ve belirli bir hacimde faj süspansiyonunun eklendiği bu katmana, eritilmiş ve 45 °C'ye soğutulmuş 2 ml %0.7 agar eklenir. Üst tabaka sertleştikten sonra fincan bir termostata yerleştirilir. Bakteriler, agarın yumuşak tabakası içinde çoğalarak faj kolonilerinin steril noktalar şeklinde açıkça görülebildiği sürekli bir opak arka plan oluşturur (Şekil 84, 2). Her koloni, bir ana faj virionunun çoğaltılmasıyla oluşturulur. Bu yöntemin kullanılması şunları sağlar: a) belirli bir materyaldeki canlı faj viryonlarının sayısını doğru bir şekilde belirlemek için kolonileri sayarak;
b) fajlardaki kalıtsal değişkenliği incelemek için karakteristik özelliklere (boyut, şeffaflık, vb.) göre.
Bakteriler üzerindeki etki spektrumuna göre fajlar ikiye ayrılır: çok değerli(liz ile ilgili bakteriler, örneğin, çok değerlikli Salmonella faj, hemen hemen tüm Salmonella'ları lize eder), monofajlar(sadece bir türün bakterilerini lize ederler, örneğin, Vi-I faj sadece tifo ateşine neden olan ajanları lize eder) ve türe özgü Bir tür içindeki bakteri varyantlarını seçici olarak parçalayan fajlar. Bu tür fajların yardımıyla, bir tür içindeki bakterilerin en ince farklılaşması, faj varyantlarına bölünmeleriyle birlikte gerçekleştirilir. Örneğin, bir dizi Vi-II faj kullanılarak, tifoya neden olan ajan 100'den fazla faj varyantına bölünür. Bakterilerin fajlara duyarlılığı, karşılık gelen reseptörlerin mevcudiyeti ile ilişkili nispeten kararlı bir özellik olduğundan, faj tiplendirmesi büyük tanısal ve epidemiyolojik öneme sahiptir.