Строение и свойства ферментов. Ферменты Ферменты ускоряют химические реакции
ОБЩАЯ ХАРАКТЕРИСТИКА ФЕРМЕНТОВ
Ферменты – биологические катализаторы.
Химическая природа ферментов. Активный центр ферментов.
Механизм ферментативного катализа.
I . Ферменты – биологические катализаторы белковой природы, способные во много раз ускорять химические реакции, протекающие в организме, но сами не входящие в состав конечных продуктов реакции.
Вещества, на которое действует фермент, называют субстратами.
Все многообразие биохимических реакций, протекающие в микроорганизмах, растениях и животных катализируется соответствующими ферментами. Велика роль ферментов в технологии пищевых продуктов. В основе производства любого пищевого продукта лежат либо биохимические (ферментативные), либо физико-химические процессы, либо эти процессы взаимосвязаны.
В отличие от неорганических катализаторов ферменты имеют свои особенности:
Скорость ферментативного катализа на несколько порядков выше (от 10 3 до 10 9), чем не биологического катализатора;
действие каждого фермента высокоспецифично, т.е. каждый фермент действует только на свой субстрат или группу родственных субстратов;
ферменты катализируют химические реакции в мягких условиях, т.е. при обычном давлении, высокой температуре (20-50С) и при значениях рН среды, в большинстве случаев близких к нейтральной.
С точки зрения локализации ферментов в клетке их подразделяют на внеклеточные и внутриклеточные.
Внеклеточные ферменты выделяются живой клеткой во внешнюю среду,внутриклеточные – находятся либо в клеточных органеллах, либо в комплексе с надмолекулярными структурами.
Особую группу ферментов составляют полиферментные комплексы, в состав которых входит ряд ферментов, катализирующих последовательные реакции превращения какого-либо субстрата. Эти комплексы локализованы во внутримолекулярных структурах таким образом, что каждый фермент располагается в непосредственной близости от фермента, катализирующего реакцию в цепи данной последовательности реакций. Благодаря такому расположению ферментов процесс диффузии субстрата и продуктов реакции сводится к минимуму.
II . Ферменты – высокомолекулярные белковые соединения.
Как и другие белки, ферменты имеют 4 уровня структуры, им присущи все физико-химические свойства белков, и лишь одна отличительная особенность – способность ускорять химические реакции. Ферменты могут быть простыми – однокомпонентными и сложными двухкомпонентными.
Однокомпонентные ферменты – построены из полипептидных цепей и при гидролизе распадаются только до аминокислот.
Двухкомпонентные ферменты – состоят из белковой части – апоформента и небелковой части – кофактора . Оба компонента в отдельности лишены ферментативной активности. Только соединившись вместе (холофермент ) они приобретают свойства, характерные для биокатализаторов. Роль кофактора может выполнять какой-либо ион (Zn 2+ , Mg 2+ , Fe 2+ , Cu 2+ , реже K + и Na +) или органическое соединение (витамины, нуклеотиды). Кофакторы органической природы называются коферментами.
Тип связи между кофактором и апоферментом может быть различным. В некоторых случаях они существуют отдельно и связываются только во время протекания реакции; в других случаях кофактор и апофермент связаны постоянно, иногда прочными, ковалентными связями.
Активный центр ферментов –это локальный участок молекулы фермента, который участвует в акте катализа. Воднокомпонентных ферментахактивный центр образуется в результате определенной ориентации аминокислотных остатков полипептидной цепи. Обычно в его формировании принимает участие небольшое количество аминокислот, в пределах 12-16. Функциональные группы этих аминокислот могут принадлежать звеньям полипептидной цепи, удаленным друг от друга. Их сближение связано с формированием третичной структуры ферменты.
В двухкомпонентных ферментах активный центр представляет собой комплекс кофактора и некоторых примыкающих к нему аминокислотных остатков.
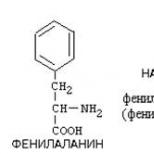
В активном центре различают контактный (якорный ) участок, функция которого – связывать субстрат, икаталитический – где происходит превращение субстрата в продукты реакции после его связывания контактным участком. В формировании этих участков принимают участие следующие функциональные группы: СООН-группы дикарбоновых аминокислот или концевые группы полипептидной цепи; имидазольная группа гистидина; ОН-группа серина;NH 2 - группа лизин и концевые группы полипептидной цепи; фенольная группа тирозина и гидрофобные остатки алифатических аминокислот.
III . Скорость любой ферментативной реакции определяетсяэнергетическим барьером , который необходимо преодолеть реагирующим молекулам. По Аррениусу, химическая реакция с точки зрения энергетики процесса описывается уравнением
N = N 0 e -(E акт /RT) ,
где N– число активных молекул;N 0 - общее число реагирующих молекул; е – основание натурального логарифма;R– газовая постоянная;T– абсолютная температура; Е акт – энергия активации.
Энергия активации – дополнительное количество энергии, необходимое для того, чтобы все молекулы преодолели энергетический барьер реакции и вступили в неё. Эта энергия представляет собой разность общей энергии реагирующих молекул и энергиивозбужденного переходного состояния. Чем больше энергия активации в реагирующей системе, тем выше энергетический барьер и тем ниже скорость реакции.
Важнейшая функция фермента – снижение энергии активации катализируемого процесса. На рис. 1 представлен график изменения энергии неферментативной (1) и ферментативной (2) реакций. Фермент снижает высоту энергетического барьера (Е акт Е акт).
Механизм ферментативного катализа во многом остается пока еще не выясненным. Однако большую роль в создании ферментативной кинетики сыграли работы М. Михаэлиса и М. Ментен, в которых было развито представление о фермент-субстратном комплексе . Образование этого комплекса и ведет к снижению энергии активации реакции.
Процесс ферментативного катализа можно условно подразделить на три стадии:
Стерическое связывание субстрата Sс активным центром фермента Е (образование фермент-субстратного комплекса ЕS).
Преобразование первичного комплекса ЕSв активированный переходный комплекс ЕS ≠ .
Отделение конечного продукта Р реакции от фермента.
Первая стадия непродолжительна по времени и зависит от концентрации субстрата и фермента в среде, от скорости диффузии субстрата к активному центру фермента. В образовании комплекса ЕSмогут участвовать в различных сочетаниях как ковалентные, координационные, ионные связи, так и менее прочные формы связей - электростатическое притяжение полярных групп, ван-дер-ваальсовы силы сцепления между неполярными участками молекул, водородные связи. Характер этих связей обусловлен химическими особенностями и субстрата, и функциональных групп, входящих в активный центр фермента.
Вторая стадия является, собственно, актом катализа, т.е. актом разрыва или образования в субстрате новых связей; она наиболее медленная и лимитирует скорость протекания химической реакции. На этой стадии и происходит снижение энергии активации ферментативной реакции, за счет образования активного переходного комплекса ЕS ≠ .
На молекулярном уровне более четкое представление о механизме действия ферментов дает теория кислотно-основного катализа. Любая реакция, идущая с разрывом ковалентных связей, предполагает участие двух противоположных по характеру электронных компонентов. Электроны разрываемой связи должны оттягиваться к электро-фильному компоненту и уходить от нуклеофильного. Реагенты, которые могли бы обусловить такую электронную перестройку - это кислота и основание. Однако в одном и том же растворе создать одновременно высокие концентрации обоих компонентов невозможно, поскольку они нейтрализуют друг друга. В белковой молекуле фермента благодарязакреплению на каталитической площадке электрофильных и нуклеофильных групп не происходит прямой реакции нейтрализации. Это, собственно, и определяет акт катализа. Находясь на определенном расстоянии друг от друга, электрофильные и нуклеофильные группы каталитического участка фермента не только связываются с реагирующими группами субстрата, но и оказывают сильное поляризующее действие на группы субстрата. К этому следует добавить возможность флуктуации зарядов в комплексе ЕS, которая создает высокую степень эффективности данной поляризации. Это и является причиной снижения величины энергии активации при ферментативном катализе.
В соответствии с теорией ковалентного катализа некоторые ферменты взаимодействуют со своими субстратами, образуя нестабильные, ковалентно связанные фермент-субстратные комплексы. Из этих комплексов в ходе последующей реакции образуются продукты реакции, причем значительно быстрее, чем в случае некатализируемых реакций.
Таким образом, третья стадия, завершающаяся образованием продуктов реакции, обеспечивается процессами, протекающими на предыдущих стадиях.
ОБЩИЕ СВОЙСТВА КАТАЛИЗАТОРОВ
1. Катализаторы сами НЕ ВЫЗЫВАЮТ химическую реакцию, а только УСКОРЯЮТ реакцию, которая протекает и без них.
2. Не влияют на энергетический итог реакции.
3. В обратимых реакциях катализаторы ускоряют как прямую, так и обратную реакцию, причем в ОДИНАКОВОЙ степени, из чего следует, что катализаторы:
а) НЕ ВЛИЯЮТ на НАПРАВЛЕННОСТЬ обратимой реакции, которая определяется только соотношением концентраций исходных веществ (субстратов) и конечных продуктов;
б) НЕ ВЛИЯЮТ на ПОЛОЖЕНИЕ РАВНОВЕСИЯ обратимой реакции, а только ускоряют его достижение.
ОСОБЕННОСТИ ФЕРМЕНТОВ КАК БИОЛОГИЧЕСКИХ КАТАЛИЗАТОРОВ
Ферменты обладают всеми общими свойствами обычных катализаторов. Но, по сравнению с обычными катализаторами, все ферменты являются белками. Поэтому они обладают особенностями, отличающими их от обычных катализаторов. Эти особенности ферментов, как биологических катализаторов, иногда называют общими свойствами ферментов . К ним относятся:
1. Высокая эффективность действия. Ферменты могут ускорять реакцию в 10 8 -10 12 раз (примеры смотрите в пособии «Ферменты», стр.7-8).
2. Высокая избирательность ферментов к субстратам (субстратная специфичность) и к типу катализируемой реакции (специфичность действия) (смотрите примеры в пособии "Ферменты", стр. 14-15);
3. Высокая чувствительность ферментов к неспецифическим физико-химическим факторам среды - температуре, рН, ионной силе раствора и т.д. (смотрите пособие "Ферменты" стр.10, 12-13);
4. Высокая чувствительность к химическим реагентам;
5. Высокая и избирательная чувствительность к физико-химическим воздействиям тех или иных химических веществ, которые благодаря этому могут взаимодействовать с ферментом, улучшая или затрудняя его работу (об активаторах и ингибиторах смотрите пособие "Ферменты", стр. 27-36).
СТРОЕНИЕ ФЕРМЕНТОВ
Субстратом (S) называют вещество, химические превращения которого в продукт (Р) катализирует фермент (Е). Тот участок поверхности молекулы фермента, который непосредственно взаимодействует с молекулой субстрата, называется АКТИВНЫМ ЦЕНТРОМ ФЕРМЕНТА.
АКТИВНЫЙ ЦЕНТР ФЕРМЕНТА
Активный центр фермента образован из остатков аминокислот, находящихся в составе различных участков полипептидной цепи или различных полипептидных цепей, пространственно сближенных. Образуется на уровне третичной структуры белка-фермента.
В его пределах различают АДСОРБЦИОННЫЙ УЧАСТОК (центр) и КАТАЛИТИЧЕСКИЙ УЧАСТОК (центр). Кроме того, вне активного центра фермента встречаются особые функциональные участки; каждый из них обозначают термином АЛЛОСТЕРИЧЕСКИЙ ЦЕНТР.
КАТАЛИТИЧЕСКИЙ ЦЕНТР - это та область (зона) активного центра фермента, которая непосредственно участвует в химических преобразованиях субстрата. Формируется он за счет радикалов двух, иногда трех аминокислот, расположенных в разных местах полипептидной цепи фермента, но пространственно сближенных между собой за счет изгибов этой цепи. Например, каталитический центр "серин-гистидиновых" ферментов формируется за счет радикалов аминокислот серина и гистидина. Если фермент является сложным белком, то в формировании каталитического центра нередко участвует простетическая группа молекулы фермента (кофермент). Коферментную функцию выполняют все водорастворимые витамины и жирорастворимый витамин K. (подробнее о коферментной функции витаминов: пособие "Ферменты", стр.11).
АДСОРБЦИОННЫЙ ЦЕНТР - это участок активного центра молекулы фермента, на котором происходит сорбция (связывание) молекулы субстрата. Он формируется одним, двумя, чаще тремя радикалами аминокислот, которые обычно расположены рядом с каталитическим центром. Главная его функция - связывание молекулы субстрата и передача этой молекулы каталитическому центру в наиболее удобном положении (для каталитического центра). Эта сорбция происходит ТОЛЬКО ЗА СЧЕТ СЛАБЫХ ТИПОВ СВЯЗЕЙ и потому ЯВЛЯЕТСЯ ОБРАТИМОЙ. По мере формирования этих связей происходит конформационная перестройка адсорбционного центра, которая приводит к более тесному сближению субстрата и активного центра фермента, более точному соответствию между их пространственными конфигурациями. Такое соответствие - не заранее "готовое", а формирующееся в ходе взаимодействия - американский ученый Кошленд положил в основу теории ИНДУЦИРОВАННОГО СООТВЕТСТВИЯ (или «НАВЕДЕННОГО» СООТВЕТСТВИЯ), которая преодолела ограниченность существовавшей ранее теории КЛЮЧА И ЗАМКА (жесткого соответствия структуры субстрата структуре адсорбционного центра).
Очевидно, что именно структура адсорбционного центра определяет субстратную специфичность фермента, то есть требования фермента к молекуле химического вещества, чтобы она могла стать для него подходящим субстратом.
Некоторые вещества, обладающие подходящими характеристиками (то есть похожие на субстрат), могут тоже связываться с адсорбционным центром фермента. Но если в их молекуле нет такой химической связи, на которую может воздействовать каталитический центр данного фермента, то химических превращений этого вещества не произойдет. Занимая активный центр фермента, такие молекулы блокируют его работу, то есть являются ОБРАТИМЫМИ ИНГИБИТОРАМИ данного фермента (обратимыми, потому что связаны с ферментом слабыми типами связей). Повышая концентрацию субстрата, их можно ВЫТЕСНИТЬ из адсорбционного центра. Поэтому такие ингибиторы называют КОНКУРЕНТНЫМИ. Они конкурируют с истинным субстратом данного фермента за обладание его адсорбционным центром.
АЛЛОСТЕРИЧЕСКИМИ ЦЕНТРАМИ называют такие участки молекулы фермента вне его активного центра, которые способны связываться СЛАБЫМИ ТИПАМИ СВЯЗЕЙ (значит - обратимо) с тем или иным веществом (лигандом). Причем такое связывание приводит к такой конформационной перестройке молекулы фермента, которая распространяется и на активный центр, облегчая, либо затрудняя (замедляя) его работу. Соответственно такие вещества называются АЛЛОСТЕРИЧЕСКИМИ АКТИВАТОРАМИ или АЛЛОСТЕРИЧЕСКИМИ ИНГИБИТОРАМИ данного фермента.
Термин "аллостерический" (то есть "имеющий иную пространственную структуру") появился в связи с тем, что эти эффекторы по своей пространственной конфигурации совсем не похожи на молекулу субстрата данного фермента (и потому не могут связываться с активным центром фермента). Было сделано заключение, что и аллостерический центр не похож по своей структуре на активный центр фермента.
Аллостерические центры найдены не у всех ферментов. Они есть у тех ферментов, работа которых может изменяться под действием гормонов, медиаторов и других биологически активных веществ. Некоторые искусственно синтезированные лекарства обладают биологической активностью потому, что их молекулы комплементарны аллостерическому центру некоторых ферментов организма.
СПЕЦИФИЧНОСТЬ ФЕРМЕНТОВ
Различают два главных вида специфичности ферментов: СУБСТРАТНУЮ СПЕЦИФИЧНОСТЬ и СПЕЦИФИЧНОСТЬ ДЕЙСТВИЯ.
СУБСТРАТНАЯ СПЕЦИФИЧНОСТЬ.
Это способность фермента катализировать превращения только одного определенного субстрата или же группы сходных по строению субстратов. Определяется структурой адсорбционного участка активного центра фермента.
Различают 3 типа субстратной специфичности:
1) АБСОЛЮТНАЯ субстратная специфичность - это способность фермента катализировать превращение только одного, строго определенного субстрата.
2) ОТНОСИТЕЛЬНАЯ субстратная специфичность - способность фермента катализировать превращения нескольких, сходных по строению, субстратов.
3) СТЕРЕОСПЕЦИФИЧНОСТЬ - способность фермента катализировать превращения определенных стереоизомеров.
Например, фермент оксидаза L-аминокислот способен окислять все аминокислоты, но относящиеся только к L-ряду. Таким образом, этот фермент обладает относительной субстратной специфичностью и стереоспецифичностью одновременно.
СПЕЦИФИЧНОСТЬ ДЕЙСТВИЯ.
Специфичность действия - это способность фермента катализировать только определенный тип химической реакции.
В соответствии со специфичностью действия все ферменты делятся на 6 классов. Классы ферментов обозначаются латинскими цифрами. Название каждого класса ферментов соответствует этой цифре.
КЛАССИФИКАЦИЯ ФЕРМЕНТОВ
I класс - ОКСИДОРЕДУКТАЗЫ.
К данному классу относятся ферменты, катализирующие окислительно-восстановительные реакции. При окислении может происходить либо отнятие водорода от окисляемого вещества, либо присоединение кислорода к окисляемому веществу. В зависимости от способа окисления различают следующие подклассы оксидоредуктаз:
1) ДЕГИДРОГЕНАЗЫ. Катализируют реакции, при которых происходит отнятие водорода от окисляемого вещества. Пример:
2) ОКСИГЕНАЗЫ. Ферменты этого подкласса катализируют включение кислорода в окисляемое вещество.
a) Монооксигеназы - включают один атом кислорода в окисляемое вещество.

б)Диоксигеназы - включают 2 атома кислорода в окисляемое вещество. Часто это сопровождается разрывом циклической структуры. По месту разрыва связи (на рисунке обозначено стрелкой) присоединяются атомы кислорода.

II класс - ТРАНСФЕРАЗЫ
Катализируют реакции переноса химических групп с молекулы одного вещества на молекулу другого вещества.
III класс - ГИДРОЛАЗЫ
Катализируют реакции разрушения химических связей с участием воды.
IV класс - ЛИАЗЫ
Катализируют реакции разрушения химических связей без участия воды.
V класс - ИЗОМЕРАЗЫ
Катализируют реакции изомерных превращений.
ИНГИБИТОРЫ ФЕРМЕНТОВ
Ингибиторы ферментов - это вещества, замедляющие ферментативные реакции.

СТРОЕНИЕ И СВОЙСТВА ФЕРМЕНТОВ
ФЕРМЕНТЫ (энзимы) - это высокоспецифичные белки, выполняющие функции биологических катализаторов . Катализатор - это вещество, которое ускоряет химическую реакцию, но само в ходе этой реакции не расходуется.
Какие условия необходимы для химического взаимодействия молекул, чтобы произошла химическая реакция?
1) Молекулы должны сблизиться (столкнуться) . Но не всякое столкновение приводит к взаимодействию.
2) Необходимо, чтобы это столкновение стало эффективным - завершилось бы химическим превращением . Обязательное условие для эффективности столкновения - чтобы запас энергии молекул в момент столкновения был не ниже энергетического уровня реакции .
ЭНЕРГЕТИЧЕСКИЙ УРОВЕНЬ РЕАКЦИИ - это запас энергии, которым должны обладать молекулы, чтобы их столкновение стало эффективным (чтобы произошла химическая реакция) . Этот запас энергии является постоянной характеристикой (константой) для каждой данной реакции.
СРЕДНИЙ ЭНЕРГЕТИЧЕСКИЙ УРОВЕНЬ МОЛЕКУЛ – это энергия, которой обладает большинство молекул системы в данный момент времени. Этасредняя величина энергетического запаса, которая характеризует совокупность данных молекул в данных конкретных условиях (температура, давление и другие). Энергетический запас молекул - это понятие статистическое (вероятностное). Молекулы постоянно находятся в тепловом движении. Поэтому энергетический запас каждой из них все время изменяется, колеблется около величины, которая и представляет собой СРЕДНИЙ ЭНЕРГЕТИЧЕСКИЙ УРОВЕНЬ МОЛЕКУЛ.
 |
В каждый момент времени наибольшая доля молекул данной совокупности обладает именно таким СРЕДНИМ запасом энергии. И чем больше отличается энергия определенной группы молекул от среднего энергетического уровня (в любую сторону), тем малочисленнее эта группа. В любой совокупности молекул ее определенная доля обладает такой энергией, которая выше среднего энергетического уровня и достаточна для протекания химической реакции.
РАЗНОСТЬ между СРЕДНИМ ЭНЕРГЕТИЧЕСКИМ УРОВНЕМ МОЛЕКУЛ и ЭНЕРГЕТИЧЕСКИМ УРОВНЕМ РЕАКЦИИ называется энергетическим барьером или ЭНЕРГИЕЙ АКТИВАЦИИ (П акт. ). Чем больше эта энергия активации, тем медленнее идет химическая реакция.
КАК УСКОРИТЬ ХИМИЧЕСКУЮ РЕАКЦИЮ? Повышение среднего энергетического уровня молекул (повышение температуры, давления и других параметров среды, которое используют на химических заводах и фабриках) НЕВОЗМОЖНО для живых организмов, которые нормально функционируют только при постоянных значениях температуры, давления и других параметров. Невозможен и другой путь - уменьшение энергии активации путем снижения энергетического уровня реакции, поскольку эта величина является постоянной характеристикой данной реакции.
Поэтому, только явление катализа (применение катализаторов) может обеспечить ускорение химических реакций в живых организмах. Рассмотрим две реакции (смотрите рисунок).
 |
В общем случае энергии активации реакций 1, 2а и 2б не совпадают между собой, и все разнообразие возможных вариантов можно разделить на две группы:
1) Еакт 2а и/или Еакт 2б БОЛЬШЕ, чем Еакт 1 .
Во всех таких случаях реакция образования вещества "АВ" с участием вещества "К" пойдет медленнее. Значит, вещество "К" является ИНГИБИТОРОМ (замедлителем) этой реакции.
2) Еакт 2а и/или Еакт 2б МЕНЬШЕ, чем Еакт 1 .
В этих случаях реакция с участием вещества "К" пойдет быстрее, чем без него. Значит, вещество "К" является КАТАЛИЗАТОРОМ (ускорителем) данной химической реакции.
КАТАЛИЗАТОР - это вещество, которое направляет реакцию по такому обходному пути, на котором энергетические барьеры ниже.
Рассмотренные реакции можно представить в виде "вертикального среза" через вершину вулкана, кратер которого имеет неодинаковые по высоте края.
 |
На этом рисунке видно, что энергия, которую надо затратить для "подъема" молекулы от среднего энергетического уровня реакции, полностью компенсируется при самостоятельном "скатывании" молекулы по склону "вулкана". При дальнейшем самопроизвольном "скатывании" до подошвы "вулкана" (то есть, до среднего энергетического уровня молекул, которые являются продуктами данной реакции). Энергия, которая при этом выделяется, называется "ЭНЕРГЕТИЧЕСКИЙ ИТОГ РЕАКЦИИ".
ЭНЕРГЕТИЧЕСКИЙ ИТОГ РЕАКЦИИ - это разность между энергетическим уровнем исходных веществ (субстратов) и энергетическим уровнем продуктов реакции.
Энергетический итог реакции не зависит от пути, по которому идет реакция (он одинаков и для реакции с участием катализатора, и для реакции без его участия). Он не зависит и от величины энергии активации - от нее зависит только скорость протекания каждого из путей этой реакции.
Это видно из УРАВНЕНИЯ АРРЕНИУСА:

Вывод из уравнения Аррениуса: так как энергия активации в этом уравнении входит в показатель степени, то даже маленькое изменение энергии активации приводит к большим изменениям скорости реакции.
Глава IV .3.
Ферменты
Обменвеществ в организме можно определить как совокупность всех химических превращений, которым подвергаются соединения, поступающие извне. Эти превращения включают все известные виды химических реакций: межмолекулярный перенос функциональных групп, гидролитическое и негидролитическое расщепления химических связей, внутримолекулярная перестройка, новообразование химических связей и окислительно - восстановительные реакции. Такие реакции протекают в организме с чрезвычайно большой скоростью только в присутствии катализаторов. Все биологические катализаторы представляют собой вещества белковой природы и носят названия ферменты (далее Ф) или энзимы (Е).
Ферменты не являются компонентами реакций, а лишь ускоряют достижение равновесия увеличивая скорость как прямого, так и обратного превращения. Ускорение реакции происходит за счет снижении энергии активации – того энергетического барьера, который отделяет одно состояние системы (исходное химическое соединение) от другого (продукт реакции).
Ферменты ускоряют самые различные реакции в организме. Так достаточно простая с точки зрения традиционной химии реакция отщепления воды от угольной кислоты с образованием СО 2 требует участия фермента, т.к. без него она протекает слишком медленно для регулирования рН крови. Благодаря каталитическому действию ферментов в организме становится возможным протекание таких реакций, которые без катализатора шли бы в сотни и тысячи раз медленнее.
Свойства ферментов
1. Влияние на скорость химической реакции: ферменты увеличивают скорость химической реакции, но сами при этом не расходуются.
Скорость реакции – это изменение концентрации компонентов реакции в единицу времени. Если она идет в прямом направлении, то пропорциональна концентрации реагирующих веществ, если в обратном – то пропорциональна концентрации продуктов реакции. Отношение скоростей прямой и обратной реакций называется константой равновесия. Ферменты не могут изменять величины константы равновесия, но состояние равновесия в присутствии ферментов наступает быстрее.
2. Специфичность действия ферментов. В клетках организма протекает 2-3 тыс. реакций, каждая из которые катализирутся определенным ферментом. Специфичность действия фермента – это способность ускорять протекание одной определенной реакции, не влияя на скорость остальных, даже очень похожих.
Различают:
Абсолютную – когда Ф катализирует только одну определенную реакцию (аргиназа – расщепление аргинина)
Относительную (групповую спец) – Ф катализирует определенный класс реакций (напр. гидролитическое расщепление) или реакции при участии определенного класса веществ.
Специфичность ферментов обусловлена их уникальной аминокислотной последовательностью, от которойзависит конформация активного центра, взаимодействующего с компонентами реакции.
Вещество, химическое превращение которого катализируется ферментом носит название субстрат ( S ) .
3. Активность ферментов – способность в разной степени ускорять скорость реакции. Активность выражают в:
1) Международных единицах активности – (МЕ) количество фермента, катализирующего превращение 1 мкМ субстрата за 1 мин.
2)Каталах (кат) – количество катализатора (фермента), способное превращать 1 моль субстрата за 1 с.
3) Удельной активности – число единиц активности (любых из вышеперечисленных) в исследуемом образце к общей массе белка в этом образце.
4) Реже используют молярную активность – количество молекул субстрата превращенных одной молекулой фермента за минуту.
Активность зависит в первую очередь от температуры . Наибольшую активность тот или иной фермент проявляет при оптимальной температуре. Для Ф живого организма это значение находится в пределах +37,0 - +39,0 ° С, в зависимости от вида животного. При понижении температуры, замедляется броуновское движение, уменьшается скорость диффузии и, следовательно, замедляется процесс образования комплекса между ферментом и компонентами реакции (субстратами). В случае повышения температуры выше +40 - +50 ° С молекула фермента, которая является белком, подвергается процессу денатурации. При этом скорость химической реакции заметно падает (рис. 4.3.1.).
Активность ферментовзависит также от рН среды . Для большинства из них существует определенное оптимальное значение рН, при котором их активность максимальна. Поскольку в клетке содержатся сотни ферментов и для каждого из них существуют свои пределы опт рН, то изменение рН это один из важных факторов регуляции ферментативной активности. Так, в результате одной химреакции при участии определенного фермента рН опт которого лежит в перделах 7.0 – 7.2 образуется продукт, который является кислотой. При этом значение рН смещается в область 5,5 – 6.0. Активность фермента резко снижается, скорость образования продуктазамедляется, но при этом активизируется другой фермент, для которого эти значения рН оптимальны и продукт первой реакции подвергается дальнейшему химическому превращению. (Еще пример про пепсин и трипсин).
Химическая природа ферментов. Строение фермента. Активный и аллостерический центры
Все ферменты это белки с молекулярной массой от 15 000 до нескольких млн Да. По химическому строению различают простые ферменты (состоят только из АК) и сложные ферменты (имеют небелковую часть или простетическую группу). Белковая часть носит название – апофермент, а небелковая, если она связана ковалентно с апоферментом, то называется кофермент, а если связь нековалентная (ионная, водородная) – кофактор . Функции простетической группы следующие: участие в акте катализа, осуществление контакта между ферментом и субстратом, стабилизация молекулы фермента в пространстве.
В роли кофактора обычно выступают неорганические вещества- ионы цинка, меди, калия, магния, кальция, железа, молибдена.
Коферменты можно рассматривать как составную часть молекулы фермента. Это органические вещества, среди которых различают: нуклеотиды (АТФ , УМФ , и пр), витамины или их производные (ТДФ – из тиамина (В 1 ), ФМН – из рибофлавина (В 2 ), коэнзим А – из пантотеновой кислоты (В 3 ), НАД и пр) и тетрапиррольные коферменты – гемы.
В процессе катализа реакции в контакт с субстратом вступает не вся молекула фермента, а определенный ее участок, который называется активным центром . Эта зона молекулы не состоит из последовательности аминокислот, а формируется при скручивании белковой молекулы в третичную структуру. Отдельные участки аминокислот сближаются между собой, образуя определенную конфигурацию активного центра. Важная особенность строения активного центра - его поверхность комплементарна поверхности субстрата, т.е. остатки АК этой зоны фермента способны вступать в химическое взаимодействие с определенными группами субстрата. Можно представить, что активный центр фермента совпадает со структурой субстрата как ключ и замок.
В активном центре различают две зоны: центр связывания , ответственный за присоединение субстрата, и каталитический центр , отвечающий за химическое превращение субстрата. В состав каталитического центра большинства ферментов входят такие АК, как Сер, Цис, Гис, Тир, Лиз. Сложные ферменты в каталитическом центре имеют кофактор или кофермент.
Помимо активного центра ряд ферментов снабжен регуляторным (аллостерическим) центром. С этой зоной фермента взаимодействуют вещества, влияющие на его каталитическую активность.
Механизм действия ферментов
Акт катализа складывается из трех последовательных этапов.
1. Образование фермент-субстратного комплекса при взаимодействии через активный центр.
2. Связывание субстрата происходит в нескольких точках активного центра, что приводит к изменению структуры субстрата, его деформации за счет изменения энергии связей в молекуле. Это вторая стадия и называется она активацией субстрата. При этом происходит определенная химическая модификация субстрата и превращение его в новый продукт или продукты.
3. В результате такого превращения новое вещество (продукт) утрачивает способность удерживаться в активном центре фермента и фермент-субстратный, вернее уже фермент-продуктный комплекс диссоциирует (распадается).
Виды каталитических реакций:
А+Е = АЕ = БЕ = Е + Б
А+Б +Е = АЕ+Б = АБЕ = АБ + Е
АБ+Е = АБЕ = А+Б+Е,где Е - энзим, А и Б - субстраты, либо продукты реакции.
Ферментативные эффекторы - вещества, изменяющие скорость ферментативного катализа и регулирующие за счет этого метаболизм. Среди них различают ингибиторы - замедляющие скорость реакции и активаторы - ускоряющие ферментативную реакцию.
В зависимости от механизма торможения реакции различают конкурентные и неконкурентные ингибиторы. Строение молекулы конкурентного ингибитора сходно со структурой субстрата и совпадает с поверхностью активного центра как ключ с замком (или почти совпадает). Степень этого сходства может даже быть выше чем с субстратом.
Если А+Е = АЕ = БЕ = Е + Б, тоИ+Е = ИЕ ¹
Концентрация способного к катализу фермента при этом снижается и скорость образование продуктов реакции резко падает (рис. 4.3.2.).

В качестве конкурентных ингибиторов выступает большое число химических веществ эндогенного и экзогенного происхождения (т.е. образующихся в организме и поступающих извне – ксенобиотики, соответственно). Эндогенные вещества являются регуляторами метаболизма и называются антиметаболитами. Многие из них используют при лечении онкологических и микробных заболеваний, тк. они ингибируют ключевые метаболичекие реакции микроорганизмов (сульфаниламиды) и опухолевых клеток. Но при избытке субстрата и малой концентрации конкурентного ингибитора его действие отменяется.
Второй вид ингибиторов - неконкурентные. Они взаимодействую с ферментом вне активного центра и избыток субстрата не влияет на их ингибирующую способность, как в случае с конкурентными ингибиторами. Эти ингибиторы взаимодействуют или с определенными группами фермента (тяжелые металлы связываются с тиоловыми группами Цис) или чаще всего регуляторным центром, что снижает связывающую способность активного центра. Собственно процесс ингибирования - это полное или частичное подавление активности фермента при сохранении его первичной и пространственной структуры.
Различают также обратимое и необратимое ингибирование. Необратимые ингибиторы инактивируют фермент, образуя с его АК или другими компонентами структуры химическую связь. Обычно это ковалентная связь с одним из участков активного центра. Такой комплекс практически недиссоциирует в физиологических условиях. В другом случае ингибитор нарушает конформационную структуру молекулы фермента - вызывает его денатурацию.
Действие обратимых ингибиторов может быть снято при переизбытке субстрата или под действием веществ, изменяющих химическую структуру ингибитора. Конкурентные и неконкурентные ингибиторы относятся в большинстве случаев к обратимым.
Помимо ингибиторов известны еще активаторы ферментативного катализа. Они:
1) защищают молекулу фермента от инактивирующих воздействий,
2) образуют с субстратом комплекс, который более активно связывается с активным центром Ф,
3) взаимодействуя с ферментом, имеющим четвертичную структуру, разъединяют его субъединицы и тем самым открывают доступ субстрату к активному центру.
Распределение ферментов в организме
Ферменты, участвующие в синтезе белков, нуклеиновых кислот и ферменты энергетического обмена присутствуют во всех клетках организма. Но клетки, которые выполняют специальные функции содержат и специальные ферменты. Так клетки островков Лангерганса в поджелудочной железе содержат ферменты, катализирующие синтез гормонов инсулина и глюкагона. Ферменты, свойственные только клеткам определенных органов называют органоспецифическими: аргиназа и урокиназа - печень, кислая фосфатаза - простата. По изменению концентрации таких ферментов в крови судят о наличии патологий в данных органах.
В клетке отдельные ферменты распределены по всей цитоплазме, другие встроены в мембраны митохондрий и эндоплазматического ретикулума, такие ферменты образуют компартменты, в которых происходят определенные, тесно связанные между собой этапы метаболизма.
Многие ферменты образуются в клетках и секретируются в анатомические полости в неактивном состоянии - это проферменты. Часто в виде проферментов образуются протеолитические ферменты (расщепляющие белки). Затем под воздействием рН или других ферментов и субстратов происходит их химическая модификация и активный центр становится доступным для субстратов.
Существуют также изоферменты - ферменты, отличающиеся по молекулярной структуре, но выполняющие одинаковую функцию.
Номенклатура и классификация ферментов
Название фермента формируется из следующих частей:
1. название субстрата с которым он взаимодействует
2. характер катализируемой реакции
3. наименование класса ферментов (но это необязательно)
4. суффикс -аза-
пируват - декарбоксил - аза,сукцинат - дегидроген - аза
Посколькууже известно порядка 3 тыс. ферментов их необходимо классифицировать. В настоящее время принята международная классификация ферментов, в основу которой положен тип катализируемой реакции. Выделяют 6 классов, которые в свою очередь делятся на ряд подклассов (в данной книге представлены только выборочно):
1. Оксидоредуктазы. Катализируют окислительно-восстановительные реакции. Делятся на 17 подклассов. Все ферменты содержат небелковую часть в виде гема или производных витаминов В 2 , В 5 . Субстрат, подвергающийся окислению выступает как донор водорода.
1.1. Дегидрогеназы отщепляют от одного субстрата водород и переносят на другие субстраты. Коферменты НАД, НАДФ, ФАД, ФМН. Они акцептируют на себе отщепленный ферментом водород превращаясь при этом в восстановленную форму (НАДН, НАДФН, ФАДН) и переносят к другому фермент-субстратному комплексу, где его и отдают.
1.2. Оксидазы - катализирует перенос водорода на кислород с образованием воды или Н 2 О 2 . Ф. Цитохромокисдаза дыхательной цепи.
RH + NAD H + O 2 = ROH + NAD + H 2 O
1.3. Монооксидазы - цитохром Р450 . По своему строению одновременно гемо- и флавопротеид. Он гидроксилирует липофильные ксенобиотики (по вышеописанному механизму).
1.4. Пероксидазы и каталазы - катализируют разложение перекисиводорода, которая образуется в ходе метаболических реакций.
1.5. Оксигеназы - катализируют реакции присоединения кислорода к субстрату.
2. Трансферазы - катализируют перенос различных радикалов от молекулы донора к молекуле акцептору.
Аа + Е + В = Еа + А + В = Е + Ва + А
2.1. Метилтрансферазы (СН 3 -).
2.2.Карбоксил- и карбамоилтрансферазы.
2.2. Ацилтрансферазы – Кофермент А (перенос ацильной группы - R -С=О).
Пример: синтез нейромедиатора ацетилхолина (см.главу "Обмен белков").
2.3. Гексозилтрансферазы- катализируют перенос гликозильных остатков.
Пример: отщепление молекулы глюкозы от гликогена под действием фосфорилазы .
2.4. Аминотрансферазы - перенос аминогрупп
R 1- CO - R 2 + R 1 - CH - NH 3 - R 2 = R 1 - CH - NH 3 - R 2 + R 1- CO - R 2
Играют важную роль в превращении АК. Общим коферментом являнтся пиридоксальфосфат.
Пример: аланинаминотрансфераза (АлАТ): пируват + глутамат = аланин + альфа-кетоглутарат (см.главу "Обмен белков").
2.5. Фосфотрансфереза (киназа) - катализируют перенос остатка фосфорной кислоты. В большинстве случает донором фосфата является АТФ. В процессе расщепления глюкозы в основном принимают участие ферменты этого класса.
Пример: Гексо (глюко)киназа .
3. Гидролазы - катализируют реакции гидролиза, т.е. расщепление веществ с присоединением по месту разрыва связи воды. К этому классу относятся преимущественно пищеварительные ферменты, они однокомпонентные (не содержат небелковой части)
R1-R2 +H 2 O = R1H + R2OH
3.1.
Эстеразы - расщепляют эфирные связи. Это большой
подкласс ферментов, катализирующих гидролиз тиоловых эфиров, фосфоэфиров.
Пример: NH
2
).
Пример: аргиназа (цикл мочевины).
4.Лиазы - катализируют реакции расщепления молекул без присоединения воды. Эти ферменты имеют небелковую часть в виде тиаминпирофосфата (В 1) и пиридоксальфосфата (В 6).
4.1. Лиазы связи С-С. Их обычно называют декарбоксилазами.
Пример: пируватдекарбоксилаза .
5.Изомеразы - катализируют реакции изомеризации.
Пример: фосфопентозоизомераза, пентозофосфатизомераза (ферменты неокислительной ветви пентозофосфатного пути).
6.Лигазы катализируют реакции синтеза более сложных веществ из простых. Такие реакции идут с затратой энергии АТФ. К названию таких ферментов прибавляют "синтетаза".
ЛИТЕРАТУРА К ГЛАВЕ IV .3.
1. Бышевский А. Ш., Терсенов О. А. Биохимия для врача // Екатеринбург: Уральский рабочий, 1994, 384 с.;
2. Кнорре Д. Г., Мызина С. Д. Биологическая химия. – М.: Высш. шк. 1998, 479 с.;
3. Филиппович Ю. Б., Егорова Т. А., Севастьянова Г. А. Практикум по общей биохимии // М.: Просвящение, 1982, 311с.;
4. Ленинджер А. Биохимия. Молекулярные основы структуры и функций клетки // М.: Мир, 1974, 956 с.;
5. Пустовалова Л.М. Практикум по биохимии // Ростов-на Дону: Феникс, 1999, 540 с.
Важнейшей функцией белков является каталитическая, ее выполняет определенный класс белков – ферменты. В организме выявлено более 2000 ферментов. Ферменты – это биологические катализаторы белковой природы, которые значительно ускоряют биохимические реакции. Так, ферментативная реакция происходит в 100-1000 раз быстрее, чем без ферментов. Многими свойствами они отличаются от катализаторов, использующихся в химии. Ферменты ускоряют реакции при обычных условиях, в отличие от химических катализаторов.
В организме человека и животных за несколько секунд происходит сложная последовательность реакций, для проведения которой с применением обычных химических катализаторов требуется продолжительное время (дни, недели или даже месяцы). В отличие от реакций без ферментов, в ферментативных не образуются побочные продукты (выход конечного продукта – почти 100 %). В процессе преобразований ферменты не разрушаются, поэтому небольшое их количество способно катализировать химические реакции большого количества веществ. Все ферменты – белки и имеют характерные для них свойства (чувствительность к изменениям pH среды, денатурация при высоких температурах и т. п.).
Ферменты по химической природе разделяют на однокомпонентные (простые) и двухкомпонентные (сложные) .
Однокомпонентные (простые) ферменты
Однокомпонентные ферменты состоят только из белков. К простым принадлежат преимущественно ферменты, которые осуществляют реакции гидролиза (пепсин, трипсин, амилаза, папаин и т. п.).
Двухкомпонентные (сложные) ферменты
В отличие от простых, сложные ферменты содержат небелковую часть – низкомолекулярный компонент. Белковая часть называется апоферментом (носителем фермента), небелковая – коферментом (активной или простетичной группой). Небелковая часть ферментов может быть представлена или органическими веществами (например, производными витаминов, НАД, НАДФ, уридиновыми, цитидиловыми нуклеотидами, флавинами), или неорганическими (например, атомами металлов – железа, магния, кобальта, меди, цинка, молибдена и т. п.).
Не все необходимые коферменты могут синтезироваться организмами и потому должны поступать с пищей. Отсутствие витаминов в пище человека и животных служит причиной потери или снижения активности тех ферментов, в состав которых они входят. В отличие от белковой части органические и неорганические коферменты очень стойкие к неблагоприятным условиям (высокой или низкой температурам, излучению и т.п.) и могут отделяться от апофермента.
Характеризуются ферменты высокой специфичностью: могут превращать лишь соответствующие субстраты и катализировать лишь определенные реакции одного типа. Определяет ее белковый компонент, но не вся его молекула, а лишь ее небольшой участок – активный центр . Структура его отвечает химическому строению веществ, которые вступают в реакцию. Для ферментов характерно пространственное соответствие между субстратом и активным центром. Они подходят друг другу, как ключ замку. Активных центров может быть несколько в одной молекуле фермента. Активный центр, то есть место соединения с другими молекулами, есть не только у ферментов, а и у некоторых других белков (гем в активных центрах миоглобина и гемоглобина). Протекают ферментативные реакции в виде последовательных этапов – от нескольких до десятков.
Активность сложных ферментов проявляется лишь тогда, когда белковая часть соединяется с небелковой. Также их активность проявляется лишь при определенных условиях: температуры, давления, pH среды и т. п. Ферменты разных организмов наиболее активны при температуре, к которой приспособлены эти существа.
Фермент-субстратный комплекс
Связи субстрата с ферментом образуют фермент-субстратный комплекс.
При этом он изменяет не только собственную конформацию, а и конформацию субстрата. Ферментативные реакции могут тормозиться собственными продуктами реакции – при накоплении продуктов скорость реакции снижается. Если продуктов реакции мало, то фермент активируется.
Вещества, проникающие в область активного центра и блокирующие каталитические группы ферментов, называются ингибиторами (от лат. inhibere – сдерживать, останавливаться). Активность ферментов снижают ионы тяжелых металлов (свинец, ртуть и т.п.).
Ферменты уменьшают энергию активации, то есть уровень энергии, необходимый для придания реакционной способности молекулам.
Энергия активации
Энергия активации – это энергия, которая расходуется на разрыв определенной связи для химического взаимодействия двух соединений. Ферменты имеют определенное расположение в клетке и организме в целом. В клетке ферменты содержатся в определенных ее частях. Многие из них связаны с мембранами клеток или отдельных органелл: митохондрий, пластид и т. п.
Биосинтез ферментов организмы способны регулировать. Это позволяет поддерживать относительно постоянный их состав при значительных изменениях условий окружающей среды и частично видоизменять ферменты в ответ на такие изменения. Действие разных биологически активных веществ–гормонов, лекарственных препаратов, стимуляторов роста растений, ядов и т. п. – заключается в том, что они могут стимулировать или подавлять тот или иной ферментативный процесс.
Некоторые ферменты принимают участие в активном транспорте веществ через мембраны.
Для названий большинства ферментов характерен суффикс -аз- . Его прибавляют к названию субстрата, с которым взаимодействует фермент. Например, гидролазы – катализируют реакции расщепления сложных соединений на мономеры за счет присоединения молекулы воды в месте разрыва химической связи молекулах белков, полисахаридов, жиров; оксидредуктазы – ускоряют окислительно-восстановительные реакции (перенесение электронов или протонов); изомеразы – способствуют внутренней молекулярной перестройке (изомеризации), преобразованию изомеров и т. п.
Ферменты
Обмен веществ в организме можно определить как совокупность всех химических превращений, которым подвергаются соединения, поступающие извне. Эти превращения включают все известные виды химических реакций: межмолекулярный перенос функциональных групп, гидролитическое и негидролитическое расщепления химических связей, внутримолекулярная перестройка, новообразование химических связей и окислительно - восстановительные реакции. Такие реакции протекают в организме с чрезвычайно большой скоростью только в присутствии катализаторов. Все биологические катализаторы представляют собой вещества белковой природы и носят названия ферменты (далее Ф) или энзимы (Е).
Ферменты не являются компонентами реакций, а лишь ускоряют достижение равновесия увеличивая скорость как прямого, так и обратного превращения. Ускорение реакции происходит за счет снижении энергии активации – того энергетического барьера, который отделяет одно состояние системы (исходное химическое соединение) от другого (продукт реакции).
Ферменты ускоряют самые различные реакции в организме. Так достаточно простая с точки зрения традиционной химии реакция отщепления воды от угольной кислоты с образованием СО 2 требует участия фермента, т.к. без него она протекает слишком медленно для регулирования рН крови. Благодаря каталитическому действию ферментов в организме становится возможным протекание таких реакций, которые без катализатора шли бы в сотни и тысячи раз медленнее.
Свойства ферментов
1. Влияние на скорость химической реакции: ферменты увеличивают скорость химической реакции, но сами при этом не расходуются.
Скорость реакции – это изменение концентрации компонентов реакции в единицу времени. Если она идет в прямом направлении, то пропорциональна концентрации реагирующих веществ, если в обратном – то пропорциональна концентрации продуктов реакции. Отношение скоростей прямой и обратной реакций называется константой равновесия. Ферменты не могут изменять величины константы равновесия, но состояние равновесия в присутствии ферментов наступает быстрее.
2. Специфичность действия ферментов. В клетках организма протекает 2-3 тыс. реакций, каждая из которые катализирутся определенным ферментом. Специфичность действия фермента – это способность ускорять протекание одной определенной реакции, не влияя на скорость остальных, даже очень похожих.
Различают:
Абсолютную – когда Ф катализирует только одну определенную реакцию (аргиназа – расщепление аргинина)
Относительную (групповую спец) – Ф катализирует определенный класс реакций (напр. гидролитическое расщепление) или реакции при участии определенного класса веществ.
Специфичность ферментов обусловлена их уникальной аминокислотной последовательностью, от которой зависит конформация активного центра, взаимодействующего с компонентами реакции.
Вещество, химическое превращение которого катализируется ферментом носит название субстрат (S) .
3. Активность ферментов – способность в разной степени ускорять скорость реакции. Активность выражают в:
1) Международных единицах активности – (МЕ) количество фермента, катализирующего превращение 1 мкМ субстрата за 1 мин.
2) Каталах (кат) – количество катализатора (фермента), способное превращать 1 моль субстрата за 1 с.
3) Удельной активности – число единиц активности (любых из вышеперечисленных) в исследуемом образце к общей массе белка в этом образце.
4) Реже используют молярную активность – количество молекул субстрата превращенных одной молекулой фермента за минуту.
Активность зависит в первую очередь от температуры . Наибольшую активность тот или иной фермент проявляет при оптимальной температуре. Для Ф живого организма это значение находится в пределах +37,0 - +39,0 °С, в зависимости от вида животного. При понижении температуры, замедляется броуновское движение, уменьшается скорость диффузии и, следовательно, замедляется процесс образования комплекса между ферментом и компонентами реакции (субстратами). В случае повышения температуры выше +40 - +50 °С молекула фермента, которая является белком, подвергается процессу денатурации. При этом скорость химической реакции заметно падает (рис. 4.3.1.).
Активность ферментов зависит также от рН среды . Для большинства из них существует определенное оптимальное значение рН, при котором их активность максимальна. Поскольку в клетке содержатся сотни ферментов и для каждого из них существуют свои пределы опт рН, то изменение рН это один из важных факторов регуляции ферментативной активности. Так, в результате одной химреакции при участии определенного фермента рН опт которого лежит в перделах 7.0 – 7.2 образуется продукт, который является кислотой. При этом значение рН смещается в область 5,5 – 6.0. Активность фермента резко снижается, скорость образования продукта замедляется, но при этом активизируется другой фермент, для которого эти значения рН оптимальны и продукт первой реакции подвергается дальнейшему химическому превращению. (Еще пример про пепсин и трипсин).